Freezing Avoidance and Freezing Tolerance in Tropical High Mountain Plants
Cold hardening and de-hardening is observed only in plants of regions with seasons characterised by temperature changes. Tropical plants are not known to harden, which corresponds to virtually invariable high temperatures throughout the year. Nevertheless, plants of the tropical high mountains (e.g. the Andes, the East African and Hawaiian volcanoes, and the high mountains of New Guinea) are frost hardy. Furthermore, their frost hardiness is maintained throughout the entire year because they are subjected to the so-called Frostwechsel climate, described as summer every day and winter every night (Hedberg 1964) (Fig. 4.24).
The regularly occurring frosts are only moderate (temperatures below -15 °C are rare and occur mostly in topographic depressions) and may last for a maximum of 12 h, and the life forms of the vegetation are adapted to cope with this recurrent temperature stress (Hedberg 1964; Beck 1994a). Adaptation to this type of climate requires mechanisms of freezing avoidance in addition to acclimative reactions. The plants are frost tolerant throughout the whole year but are not tolerant of an uninterrupted frost period.
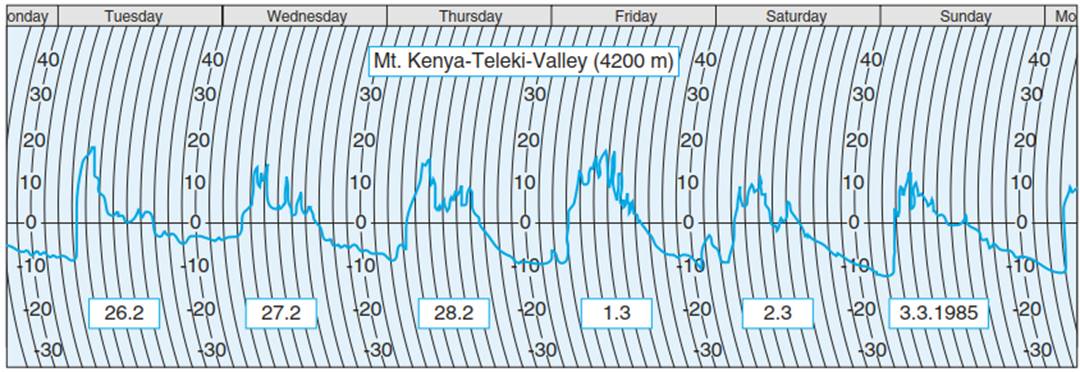
Fig. 4.24. Temperature courses in a tropical “Frostwechselklima” recorded on Mt Kenya at 4200 m altitude during the dry season. The temperature sensor was about 20 cm above the soil surface
Several types of response to sub-freezing temperatures can be recognised in the five common functional plant types of the tropical high alpine flora: grass tussocks, sclerophyllous shrubs, acaulescent rosette plants, cushion plants and the spectacular giant rosette plants. All rely on the principle of daily changes between the nocturnal frost and the “summer” temperatures during the day, caused by the intense tropical sunshine. Mechanisms to overcome the different primary and secondary abiotic stress factors (climate, high light intensities and ultraviolet radiation, seasonal shortage of water during the dry season, low partial pressure of CO2 and oxygen) imposed on the tropical alpine plants by their environment have been discussed in some detail, but the molecular level has hardly been touched upon (Beck 2011).
Ecophysiological data show that separate investigation of the impact of one abiotic stressor on these plants cannot sufficiently explain the real situation, and a more holistic consideration of their performance is required, as claimed by Gusta and Wisniewski (2013), for the understanding of plant cold hardiness. The example of the “giant rosette plant” life form (Fig. 4.25) shows that more than one “strategy” may be adopted by a plant individual—namely, freezing tolerance as well as freezing avoidance by permanent insulation (Fig. 4.25) or nocturnal insulation, thermal buffering by liquids, or supercooling.
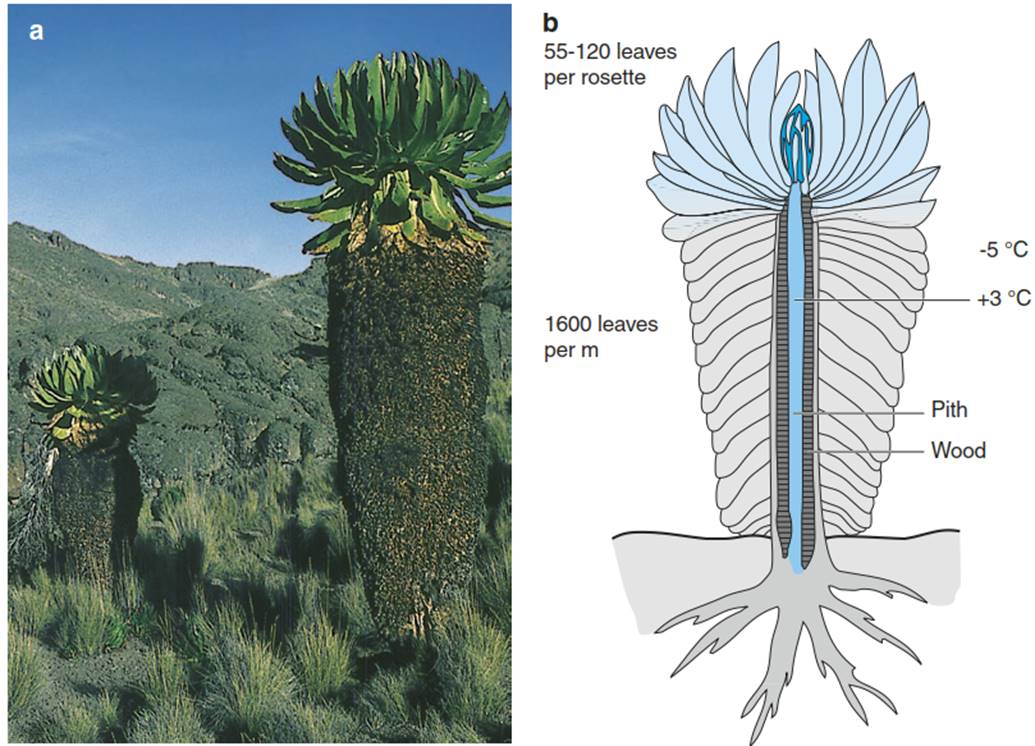
Fig. 4.25. Young specimens of the giant rosette plant Senecio (Dendrosenecio) keniodendron in the Afro-alpine zone of Mt Kenya (at 3500-4800 m). These plants carry a single terminal leaf rosette of up to 1 m diameter with around 100 big leaves surrounding the terminal leaf bud (dark blue in the sketch) in the centre of the rosette. At least the first half of the leaf development takes place in that cone-shaped leaf bud, from which the leaves separate after greening. After senescence, the dead, now outer leaves of the rosette remain attached to the stem and the resulting leaf cloak of shrivelled leaves delays nocturnal cooling of the stem until the tropical sun warms it up again.
Maintenance of the stem temperature well above the freezing point is important, as a large share of the water required by the leaves is supplied by the living pith cells, while the amount of water delivered by the extremely narrow vessels of the weakly developed xylem is inferior. Water transport via living cells depends strongly on the temperature. A similar “strategy” is known from the corresponding plant life form, Espeletia, in the tropical Andes, where the “cloak” of dead leaves has inspired the plant’s popular name, frailejones (“thick monks”). Interestingly, the cloak is densely rooted by adventitious roots from the stem, which absorb water and nutrients from the decaying leaf material. (Beck 1994b)
Each of these “strategies” may be considered to meet a special physiological requirement. While the stems, inflorescences and roots follow the “strategy” of freezing avoidance, the leaves of the African Senecio and Lobelia are freezing tolerant and are stiffly frozen in the morning before sunrise. Extracellular freezing of cellular water has been documented, concomitant with freeze cytorrhysis of mesophyll cells or intravacuolar emergence of gas bubbles, which disappear upon thawing in the hypodermis (Beck 1994a).
In nature the mesophyll cells produce sucrose as a cryoprotectant, and only a minor accumulation of starch can be detected in the chloroplasts. While the mature leaves of the giant rosette freeze every night, the young leaves in the central leaf bud, despite being freezing resistant as well, are protected from freezing through the formation of a so-called night bud (Hedberg 1964) by the outer rosette leaves. Since extracellular crystallisation of cell water results in a loss of cell turgor, growth of the young leaves would be interrupted every night.
The night bud prevents nocturnal sub-zero temperatures and concomitant freezing of the meristematic tissue, thus enabling uninterrupted growth. A question still to be solved is how propagation of ice from the frozen leaves to the other parts of the plants is prevented. Fast spreading of ice has been observed by infrared differential thermal analysis in dicotyledonous alpine plants, starting from the xylem vessels and spreading into the rest of the plants (Hacker and Neuner 2007, 2008).
In the case of Senecio, ice formation is confined to the leaves and cannot be triggered from the xylem, because of the existence of a kind of “thermal ice barrier” (Hacker et al. 2011). Interestingly, the leaves of the corresponding giant rosette plants of the neotropical Andes (Espeletia spp.) are reported to supercool to about 8-12 °C below the ambient temperature and to be killed by intracellular ice formation (Goldstein et al. 1984, 1985).
Date added: 2025-01-18; views: 533;