Evolution of C4 Photosynthesis
C4 photosynthesis is characterised by a suite of distinct anatomical, morphological, physiological and biochemical features. Therefore, it is, at first sight, surprising that C4 photosynthesis has evolved independently so many times from ancestral C3 photosynthesis. Owing to a large body of research work, we can today describe C4 photosynthesis as an excellent example of how the evolution of a key adaptation can be understood in an ecological context (Christin and Osborne 2014).
Conceptual models have been proposed as to how C4 photosynthesis developed (Fig. 6.22). Evolution along this path is plausible because for every step, distinct selective advantages can be inferred and factors priming C3 lineages for the evolution of these steps can be identified. Moreover, some of the steps can be seen in extant C3-C4 intermediate species (e.g. in the genus Flaveria), which represent different stages on the evolutionary trajectory towards C4 photosynthesis.
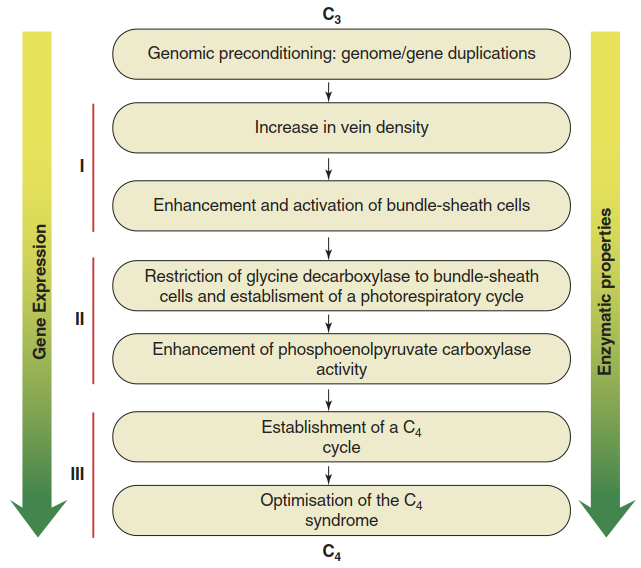
Fig. 6.22. Model of the stepwise evolution of C4 photosynthesis from C3 photosynthesis (Gowik and Westhoff 2011); see also a similar model in Sage et al. (2012)
All enzymes employed in C4 photosynthesis are of ancient bacterial origin. A first preconditioning step towards C4 photosynthesis was the duplication of respective enzyme-coding genes or of whole genomes. The existence of more than one gene copy enabled diversification with respect to localisation, timing and strength of expression. This occurred predominantly via changes in promoter sequences. Furthermore, kinetic properties could change through alterations in coding sequences (right arrow in Fig. 6.22). For example, PEP carboxylases of C4 plants are less inhibited by malate.
This is functionally important, as CO2 fixation by PEP carboxylases has to function in the presence of high malate concentrations (Fig. 6.21). The PEP carboxylase genes in C4 plants show signs of strong positive selection in their sequences, meaning there is evidence that amino acid changes affecting the kinetic properties of the encoded enzymes were selected during the evolution of C4 photosynthesis.
The efficient exchange of metabolites between mesophyll and bundle sheath cells is promoted by high vein density, as this reduces the distances between the two cell types. The high vein density typical of many C4 lineages offers advantages for C3 plants too. For example, there are more pathways for water transport into and through the leaves. This could improve drought tolerance and photosynthetic rates in arid, high-light environments.
The higher proportion of leaf volume occupied by bundle sheath cells with only a few chloroplasts and therefore low photosynthetic capacity could have exerted selective pressure to increase the number of chloroplasts and other organelles in these cells. Related to the higher metabolic activity of bundle sheath cells in C4 plants is the photorespiratory CO2 pump found in extant C3-C4 intermediates. Restriction of the glycine decarboxylase activity (for details of the photorespiration pathway, see plant biochemistry or plant physiology textbooks) to mitochondria of bundle sheath cells forces the processing of all photorespiratory glycine in these cells (sometimes this is referred to as the “C2 cycle”).
The decarboxylation releases CO2 at a site more distant from the leaf surface, thus improving the chances for refixation by RubisCO, whose oxygenation activity is suppressed by the extra CO2. Molecularly the photorespiratory CO2 pump can arise easily. It takes only two genes encoding a subunit of the glycine decarboxylase complex with expression restricted to either mesophyll or bundle sheath cells by the right cis elements in the promoters. Loss of function of the mesophyll- expressed version would then result in glycine decarboxylase activity only in the bundle sheath cells and thereby establish the CO2 pump.
Conversely, the levels of carbonic anhydrase and PEP carboxylase had to massively increase in the cytosol of the mesophyll cells. The C4 cycle is then completed through the spatial separation of the two carboxylase reactions, PEP carboxylase in the mesophyll and RubisCO in the bundle sheath cells. In the course of C4 cycle optimisation, many other metabolic changes have evolved that can mostly be explained by changes in transcriptional regulation too. A recent modelling of the biochemical fitness landscape between C3 and C4 photosynthesis—based on kinetic parameters of enzymes, gas exchange rates, etc.— demonstrated that indeed every step along the different evolutionary trajectories from C3 to C4 photosynthesis is associated with a fitness gain (Heckmann et al. 2013). This leaves the question as to why not all angiosperm lineages have evolved C4 photosynthesis. A possible explanation could be that certain potentiating factors such as high vein density are not present in all lineages.
Date added: 2025-01-27; views: 696;