Photosynthesis Variants with Improved Water Use Efficiency. C4 Photosynthesis
Plants are dependent on the uptake of CO2 through stomata, which inevitably results in evaporation of H2O. Two major variants of photosynthesis have evolved that reduce the amount of water vapour lost per unit of carbon fixed. In other words, they increase the water use effi- ciency—that is, the ratio of dry weight gained (= growth) to water lost. In regular C3 photosynthesis, about 500 g of H2O is spent per gram of carbon assimilated. The cost is reduced to about 250 g of H2O per gram of carbon in plants displaying C4 photosynthesis and to only 50-100 g of H2O in CAM plants (Table 6.3).
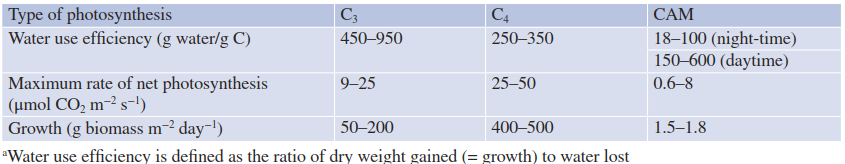
Table 6.3. Water use efficiency, photosynthesis and biomass production of C3, C4 and CAM plants. Crassulacean acid metabolism (CAM) plants are superior to other photosynthetic types in their water use efficiencya, but their photosynthetic rates and growth rates are much lower (Lüttge et al. 1994)
Both mechanisms share a first fixation of CO2 by the enzyme phosphoenolpyruvate carboxylase (PEP carboxylase) prior to a second ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO)-dependent fixation of CO2 after its release through decarboxylation of the storage molecule malate. The competitive advantage of higher water use efficiency is evident from the fact that plants showing these photosynthesis variants are predominantly found in arid and dry regions of the world.
C4 Photosynthesis. In every plant, CO2 is fixed by RubisCO. This enzyme, which represents by far the most abundant protein on Earth, catalyses the first reaction of the Calvin cycle between an activated pentose phosphate, ribulose-1,5-bisphosphate and CO2 (the carboxylase activity), yielding two molecules of 3-phosphoglycerate—a molecule with three C atoms (hence the term C3 photosynthesis). In conditions of a vast excess of O2 versus CO2, RubisCO also catalyses the reaction of ribulose-1,5-bisphosphate with O2 (the oxygenase activity), yielding one molecule of 3-phosphoglycerate and one molecule of 2-phosphoglycolate.
This side reaction poses a problem, as 2-phosphoglycolate is a useless toxic metabolite that should not accumulate. 2-Phosphoglycolate is converted to 3-phosphoglycerate via photorespiration, a pathway that requires the metabolic activity of three organelles—namely, peroxisomes and mitochondria, besides chloroplasts where 2-phosphoglycolate is produced (see plant physiology and plant biochemistry textbooks). Of the carbon in 2-phosphoglycolate, 75% is returned to the Calvin cycle as 3-phosphoglycerate.
The efficiency of ribulose-1,5-bisphosphate carboxylation is about 100-fold higher than that of the oxygenation. Thus, only gradually with the accumulation of oxygen in the atmosphere did this side reaction of RubisCO become relevant. RubisCO evolved in an atmosphere that was essentially devoid of molecular oxygen.
Furthermore, it is ecologically important that the carboxylation to oxygenation ratio of RubisCO is influenced by temperature. With higher temperature the oxygenase activity becomes more relevant because RubisCO specificity decreases. Therefore, the need for photorespiration, which negatively affects photosynthetic efficiency in plants with C3 photosynthesis, grows with increasing temperatures.
At a time about 30 million years ago, when atmospheric CO2 reached a critically low level, C4 photosynthesis arose. It has evolved independently many times since then (>60 times according to current counts) in multiple plant families. Through a series of anatomical and biochemical modifications, C4 photosynthesis achieves a higher concentration of CO2 in the vicinity of RubisCO, thereby effectively suppressing the oxygenase activity. This results in higher photosynthetic efficiency under conditions that promote photorespiration (low CO2, high temperature).
Typically, two consecutive fixations of CO2 occur in separate cell types. In mesophyll cells, which in C4 plants often surround the vascular bundles (including the bundle sheath cells) in a circular arrangement called the Kranz anatomy, CO2 is fixed by PEP carboxylase and not by RubisCO, as in C3 plants. In C4 plants, RubisCO activity is restricted to the bundle sheath cells which, in contrast to C3 plants, are more prominent and contain chloroplasts. Fixation of CO2 as bicarbonate by PEP carboxylase (which does not catalyse a reaction with O2) produces a C4 acid, which is then shuttled into the bundle sheath cells where decarboxylation releases CO2. This CO2 is utilised by RubisCO in the Calvin cycle (Fig. 6.21).
Concentration of CO2 in the vicinity of RubisCO by primary CO2 fixation via PEP carboxylase and shuttling of carbon into bundle sheath cells can be seen as the core of C4 photosynthesis. Variations between plant species exist with respect to the type of C4 acid (e.g. malate or aspartate) and the nature of the decarboxylating enzymes (malic enzyme in Fig. 6.21).
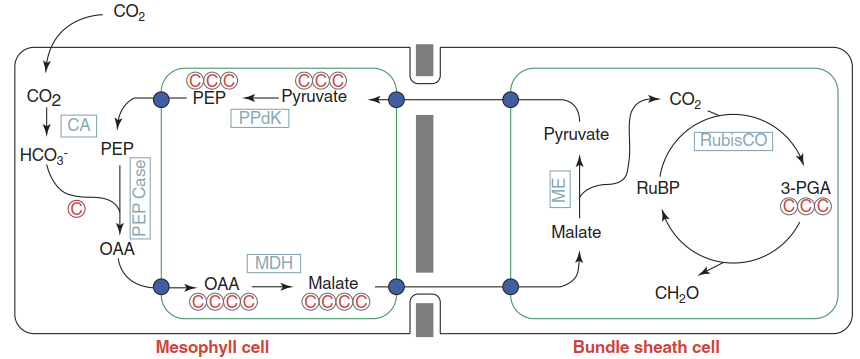
Fig. 6.21. The C4 photosynthesis core mechanism. The green boxes represent the chloroplasts. Blue dots represent active transport steps. CA carbonic anhydrase, MDHmalate dehydrogenase, ME malic enzyme, PEPCase phosphoenolpyruvate carboxylase, PPdK pyruvate orthophosphate dikinase, PEP phosphoenolpyruvate, OAA oxaloacetate, RuBP ribulose-1,5-bisphosphate, 3-PGA 3-phosphoglycerate. (Modified from Langdale (2011))
The CO2 pumping allows RubisCO to operate at near substrate saturation. This offers the potential for very high photosynthetic rates. Furthermore, it enables C4 plants to reduce stomatal conductance, which decreases water loss and improves water use efficiency. An additional advantage is the lower nitrogen requirement of C4 plants. Much less investment in RubisCO protein is needed. Higher efficiency of photosynthesis and lower demand for water explain why, today, many biomes—especially in the tropics and subtropics (such as the African savannas)—are dominated by C4 plants. Of today’s vascular plant species, 3% use C4 photosynthesis.
They account for about 25% of total terrestrial photosynthesis. Nevertheless, the distribution of C4 plants clearly shows that by no means all biomes favour C4 photosynthesis. The primary CO2 fixation consumes extra metabolic energy because the substrate PEP has to be regenerated from pyruvate, a reaction that consumes adenosine triphosphate (ATP). Thus, C4 plants have lower quantum use efficiency than C3 plants under conditions where photorespiration is low (low light, low temperature).
Date added: 2025-01-27; views: 514;