The Common Sym Pathway
Establishment of both a mycorrhiza and nitrogenfixing symbiosis has to occur for every single plant individually. It requires mutual recognition by the partners through chemical communication. Plant-derived signals perceived by the microorganisms stimulate processes that facilitate formation of the symbiosis. Conversely, signals released by the microbial partner trigger far-reaching developmental changes in the host plant that make entry of the fungal hyphae or the bacteria possible. The command over the process of symbiosis establishment lies entirely on the plant side, as the host has to actively allow infection by the symbiotic partner (Oldroyd 2013).
Plants growing in an environment with an ample supply of phosphate do not engage in mycorrhizae, and legumes provided with enough nitrate and/or ammonia do not form nodules. This illustrates why the establishment of these symbioses can be regarded as an acclimative response to nutrient scarcity.
Flavonoids released by plant roots stimulate rhizobia to synthesise nod factors. These are lipo-chito-oligosaccharides, consisting of a chitin oligomer backbone with several modifications. Nod factors are perceived by plant roots and trigger many changes in the host that enable colonisation by the bacteria. First, root hairs curl and thereby build a protected environment for the rhizobia. Following a few bacterial cell division cycles, an infection thread forms inside the root hair. Rhizobia enter the plant root through these threads. Concomitantly, nodule organogenesis occurs in the root cortex. The cell cycle is stimulated, and newly dividing cells develop into the nodule (Fig. 7.23).
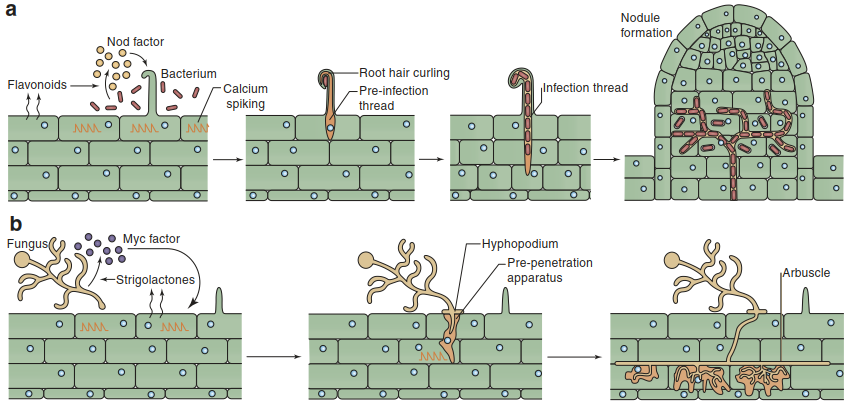
Fig. 7.23. Establishment of rhizobial and mycorrhizal root colonisation. The symbiotic partners recognise each other through the exchange of signals. Upon recognition the plant host undergoes massive developmental changes to allow colonisation. (Oldroyd 2013)
The plant signals that stimulate spore germination and hyphal branching of endomycorrhizal fungi have been identified as strigolactones (Akiyama et al. 2005)—that is, root-synthesised phytohormones involved in controlling shoot branching. Myc factors are the signals on the fungal side. They are chemically similar to nod factors in that they are lipo-chito-oligosaccharides as well (Maillet et al. 2011). Upon recognition of myc factors, plant roots undergo massive changes to allow penetration of hyphae towards the root cortex. A pre-penetration apparatus is formed in rhizodermal cells. It represents a transcellular “tunnel”, which is then used by the hyphae to grow into the root cortex (Parniske 2008). In the cortex the hyphae grow intercellularly before they form arbuscules within cells in the inner cortex (Fig. 7.23).
In spite of the fundamental differences between the symbiotic microorganisms and the plant developmental changes allowing colonisation of the host, the recognition and early signal transduction events in host cells proceed through a pathway shared by arbuscular mycorrhizae and the Fabaceae-rhizobia symbiosis: the common sym pathway. Molecular analysis of symbiotic interactions in the legume model systems has revealed that the symbiosis pathway enabling the formation of mycorrhizal associations in most plant species was, much later in evolution (around 150 million years ago), recruited by legumes for the establishment of biological nitrogen fixation. Research using mutants of M. truncatula and L. japonicus defined several genes that are essential for both mycorrhizae and root nodule symbiosis.
A lack of any of these genes (e.g. the SYMRK gene) abolishes the early responses of plant roots to the microbial signals (Stracke et al. 2002). The chemically very similar nod and myc factors are perceived by closely related receptor-like kinases residing in the plasma membrane of root cells (Fig. 7.24). The receptors and co-receptors, whose exact functions with regard to the recognition of specific ligands are not resolved yet, share LysM domains typical for proteins binding chitin fragments. Early signal transduction event in root cells stimulated by symbiosis signals are Ca oscillations in the nucleus.
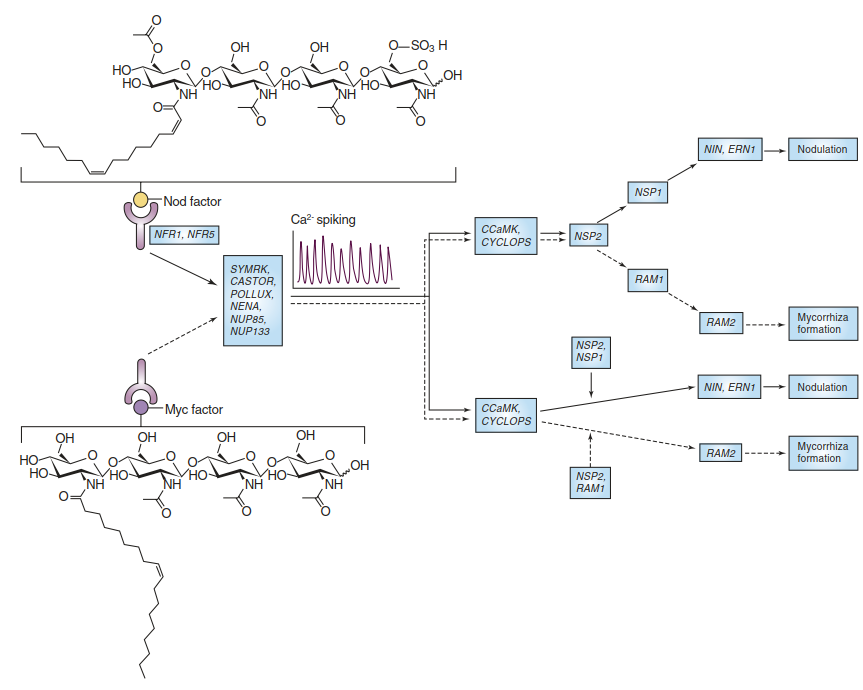
Fig. 7.24. The common sym pathway. Chemically similar bacterial and fungal signals—nod factors and myc factors, respectively—are perceived by similar receptors in the plant host cells. Downstream signal transduction initiating the developmental changes on the host side that allow establishment of either an arbuscular mycorrhiza or nodulation involves Ca oscillations triggered by the same
proteins. The Ca signal is perceived by a calcium- and calmodulin-dependent serine/threonine protein kinase (CCaMK). CYCLOPS is a protein interacting with CCaMK. Further transmission of the signal proceeds via several different transcription factors (e.g. NSP1, NSP2, RAM1) and results in either nodulation or mycorrhizal symbiosis. (Oldroyd 2013)
Components that have been identified are ion channels of the nuclear envelope (CASTOR and POLLUX) required for the induction of the calcium oscillations, and components of the nuclear pore (Oldroyd 2013). Perception of the Ca signal is mediated by Ca- and calmodulin-dependent kinases (CCaMK), which interact with another common sym pathway component of unknown molecular function, CYCLOPS (Fig. 7.24). Even among the transcription factors downstream from the Ca signalling, overlaps in functional roles between mycorrhizal establishment and nodulation exist.
The common sym pathway exists in mosses and lycophytes, illustrating once more how ancient mycorrhizal associations are in terrestrial plants. Apparently, rather subtle changes in the genomes of legumes enabled them to establish a second symbiosis much later in evolution, this time with rhizobia, by principally employing the same basic mechanisms. It has been shown that many nodulation genes can be replaced by orthologous genes from plant species that are not able to form nodules, indicating that no fundamentally new innovations were required for the legume-rhizobia symbiosis.
Nonetheless, a legume root initiates nodule formation in response to nod factors and prepares for penetration of fungal hyphae in response to myc factors. The specificity of the recognition and signalling processes is presumably conferred by specific functions of particular receptor-like kinases perceiving the signals and by specific transcriptional regulators.
Date added: 2025-01-27; views: 584;