Modulation of Nutrient Uptake in Response to Deficiency
Plants need to be able to respond and acclimate to strongly fluctuating external nutrient availability. The major targets of acclimation are nutrient uptake capacities and the root architecture. Generally relevant factors are the external supply of a particular nutrient and the internal status. Both can in principle be sensed and translated into a response. A second distinction is that between a local response (i.e. a cell or a tissue perceives a problem and triggers countermeasures) and a systemic response (i.e. leaves or another organ distant from the site of nutrient acquisition monitor the nutritional status and send a signal down to the root, where uptake is modulated appropriately).
A basal feature of plant nutrient acquisition is the switching from low-affinity uptake to high- affinity uptake when external concentrations drop below certain thresholds (Fig. 7.12). The activities of the different systems often correlate quite strongly with the expression levels of the genes encoding the transporters. Thus, transcriptional regulation is one key to the adjustment of uptake capacities. While low-affinity systems tend to be expressed constitutively, many high-affinity transporter genes (e.g. NRT2, AMT1, Pht1, KUP/HAK) have been shown to be up-regulated upon nutrient deficiency (Miller et al. 2009).
Another aspect of transcriptional control is activation in the presence of a substrate and repression by an end product. Nitrate serves as an important signal for nutrient availability and elicits the transcription of nitrate uptake transporter genes. Conversely, glutamine—as the product of ammonium transfer onto glutamate during nitrate assimilation—represents a negative feedback signal for nitrate uptake.
The P deficiency response is a relatively well- understood example of systemic signalling. Studies on A. thaliana and rice mutants with deregulated Pi acquisition (i.e. with low shoot P levels even when external supply is high) or with excessive P accumulation regardless of P status led to the identification of a systemic regulation of P uptake which, in its principal form, is present in many plants. PHO2 is a regulatory protein crucial for the down-regulation of Pht1-dependent high-affinity uptake. pho2 mutants over-accumulate P in their shoots. The PHO2 messenger RNA (mRNA) carries target sites for microRNA (miRNA) 399.
This miRNA represents a phloem- mobile signal of P status in the leaves. Low P levels stimulate miRNA399 synthesis. In roots, miRNA399 then suppresses PHO2 expression, which in turn results in higher expression of Pht1 transporters and thus higher transport capacity (Fig. 7.15). The abundance of miRNA399 is under the control of a transcription factor, Phr1, which controls many P deficiency responses.
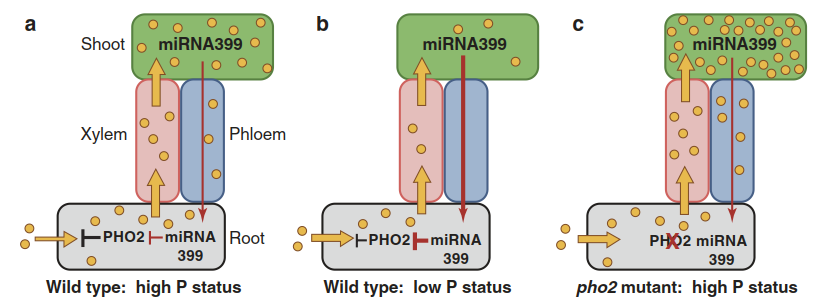
Fig. 7.15. The phosphate nutritional status of the shoot is signalled to the root by a microRNA. PHO2 is a protein that controls protein stability. When phosphate (yellow dots) supply to the shoot is sufficient a, phosphate uptake transporters (the Pht1 family) are down-regulated in a PHO2-dependent manner. However, when the shoot phosphate status is low b, microRNA (miRNA) 399 is expressed more strongly in leaf cells, travels via the phloem to the root and causes the degradation of PHO2 messenger RNA. This in turn results in higher phosphate transporter expression and hence uptake. In the pho2 mutant c, phosphate over-accumulates to toxic levels in the shoots because phosphate uptake by roots is not controlled any more. (Modified from Buchanan et al. (2015))
Locally controlled transcription and systemically controlled transcription do not fully explain the plasticity of nutrient uptake. The responsible transporters are also regulated post- translationally. The attribution of low- and high- affinity uptake to distinct classes of transporters is in fact not without exceptions. CHL1/NRT1.1, the first nitrate transporter identified in plants, can switch between a low-affinity state and a high-affinity state. In response to low external nitrate concentrations, a threonine residue is phosphorylated by a kinase (Fig. 7.16). Similar affinity switching is known to occur in high- affinity K+ uptake transporters in the HAK/KUP family. The K+ channel AKT1 is activated by phosphorylation when external K+ is low.
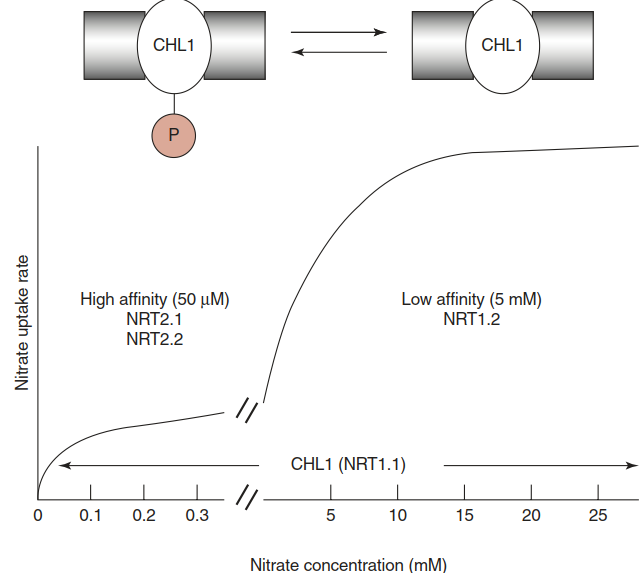
Fig. 7.16. Switching of the nitrate uptake transporter CHL1/ NRT1.1 by phosphorylation from a low-affinity status to a high-affinity status (Tsay et al. 2007)
Ammonium is a nutrient that can be toxic to cells and therefore should not accumulate. Thus, uptake of ammonium needs to be controlled by a mechanism that allows rapid shut-off if a critical ammonium concentration is exceeded. AMT proteins are oligomeric proteins. Their C-terminal domain functions as an allosteric regulator of activity, which is controlled by phosphorylation.
Also, transporter activity can cause problems when the substrate specificity is not very high. This is illustrated by the Fe(II) uptake system IRT1. IRT1 expression is transcriptionally up- regulated when the plant’s Fe status is low. Because IRT1 mediates entry also of Zn, Ni and Cd ions, among others, Fe deficiency can lead to supraoptimal uptake of these metal ions, resulting in toxicity. In the case of IRT1, the tight control through the Fe status and rapid shutdown when enough Fe is present intracellularly is brought about by cycling of the transporter within the endomembrane system. The residence time in the plasma membrane and protein stability respond immediately to Fe via post-translational mechanisms (Brumbarova et al. 2015), again enabling rapid inactivation of uptake capacity.
Date added: 2025-01-27; views: 454;