Plasticity of Root Architecture and Responses to Nutrient Deficiency
Roots are the plant organs with the highest developmental plasticity. This plasticity helps roots to efficiently explore and exploit their heterogeneous soil habitat. Uptake of water and nutrients occurs predominantly near the root tips. Continuous root growth is required to access fresh and often immobile nutrient sources.
The size of the root surface that is competent to take up water and nutrients determines the capacity of a plant individual to acquire these resources. For a given species or even for a given genotype, the spatial arrangement of roots and root systems (i.e. the root architecture) can be highly variable. It is mostly determined by:
- The number and lengths of root axes, including the length of the primary root
- The position, angle, density and growth of lateral roots
- The density and elongation of root hairs
New cells arise through cell division in the root meristem near the root tip. The root tip itself is protected by a root cap. In many species the root cap releases cells (border cells) that influence the rhizosphere—for example, by enhancing nutrient availability. Cells formed in the meristem differentiate into the epidermis, the cortex, the endodermis and the stele with its pericycle, xylem and phloem. Lateral roots are initiated in the pericycle; adventitious roots are initiated in the stem. Root hairs, which are particularly important for increasing the root surface, arise through the tip growth of designated epidermal cells, the trichoblasts.
Root architecture plasticity is governed by a variety of processes that occur in response to either the nutrient availability in the soil or the nutritional status of the plant (Fig. 7.18). Responses to nutrient availability are predominantly those triggered by the presence of nutrients. Responses to the nutritional status are triggered by nutrient deficiency. Detailed investigations have revealed clear nutrient specificity of the root architecture modulation. For instance, the strong inhibition of primary root growth and the stimulation of root hair development and elongation in phosphate-deficient plants are very well documented.
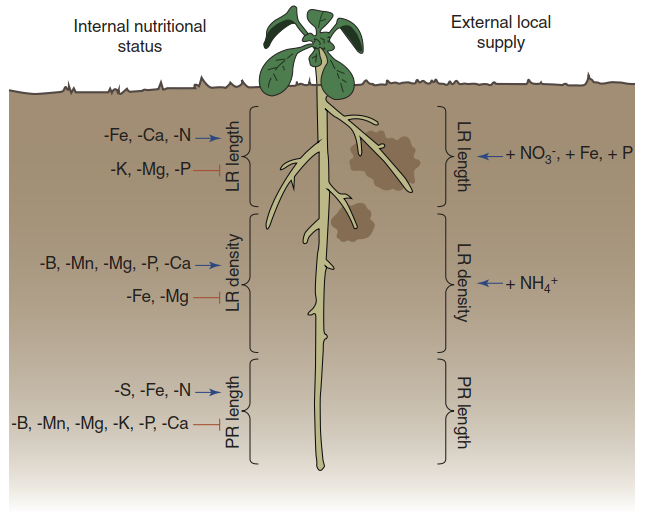
Fig. 7.18. Root developmental processes and nutrient availability responses. Blue symbols indicate stimulation, red symbols indicate inhibition. Minus (-) means deficiency of an element, plus (+) availability of an element. The root of a typical dicotyledonous plant is shown as an example. LR lateral root, PR primary root
Conversely, phosphate availability stimulates growth of lateral roots. Phosphate resources tend to be concentrated in the topsoil. The root architecture responses enhance exploitation of this soil layer. Perhaps the most striking response to phosphate deficiency is the formation of cluster roots—for instance, in a range of plants native to nutrient-poor soils in Australia (Lambers et al. 2015) (Fig. 7.19).
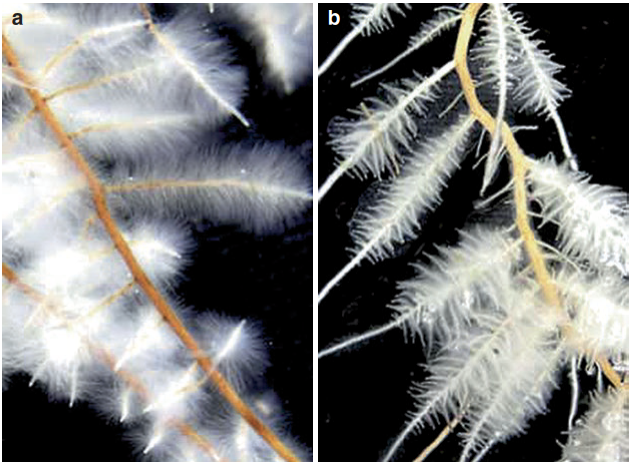
Fig. 7.19. Cluster roots of Proteaceae
Stimulation of lateral root growth by a lack of adequate nitrogen or Ca supply represents a foraging behaviour of roots. This strategy is physiologically limited inasmuch as a minimum nutrient supply has to be available to sustain the foraging (Giehl et al. 2014). When a nutrient-rich patch of soil is found, the growth of lateral roots is further stimulated. This has been shown for nitrate, phosphate and Fe (Fig. 7.18). In many experiments with various plant species, localized supply of nitrate in the substrate triggered growth of lateral roots located near or in the nutrient patch. In contrast, the systemic response of the lateral roots as a whole to conditions of ample nitrogen supply is a reduction in the growth rate.
Less investment in root biomass is needed, thus more resources can be channelled into shoot growth. The local response enables rapid exploitation of a nutrient resource and thereby gives a plant a distinct competitive advantage over its neighbours. It is dependent on the perception of nitrate as a signal and not a mere consequence of nutrient supply. Generally, phosphate and nitrogen supply elicit the strongest responses of the root system development (Osmont et al. 2007). For many root architecture responses, however, it remains to be determined whether they represent strategies to mitigate the deficiency or symptoms of either the nutrient deficiency stress or the availability of nutrients to one part of the root but not another.
Like all plant developmental processes, the formation of the root system is controlled by the delicate, fine-tuned interaction of hormones (for detailed descriptions, see plant physiology and plant molecular biology textbooks). Key events are cell cycle activity, cell differentiation and cell elongation. The interplay between auxin and cytokinin in controlling root architecture is well documented. One important aspect is the balance between cell division and cell differentiation in the root meristem (Petricka et al. 2012). An auxin gradient establishes in the root and controls stem cell maintenance, cell proliferation and differentiation. Cytokinin, on the other hand, antagonistically inhibits cell division and reduces the size of the root meristem.
Emergence of lateral roots originates from pericycle cells adjacent to the xylem poles. Auxin triggers the formation of lateral roots. The sensing of phosphate deficiency, for instance, may sensitise pericycle cells to auxin, possibly through the up-regulation of auxin receptor expression. Overall, the exact mechanisms underlying the nutrient status-dependent modulation of root architecture by the hormonal network are not well understood yet.
Sensing of Nutrient Availability and Nutrient Status. Regulation of nutrient acquisition and modulation of root system architecture require accurate sensing of external nutrient availability and internal nutritional status. The latter is monitored both locally and systemically. The respective sensing then has to be translated into transcriptional changes (for instance, the up-regulation of genes encoding high-affinity transporters), post-translational control of transporters or regulatory proteins, and changes in the concentrations and distribution of hormones.
As alluded to in previous chapters, the molecular understanding of the primary sensing of environmental parameters generally lags behind the insight into the downstream signal transduction events. This is no different for the sensing of nutrient availability in the rhizosphere or the monitoring of the nutritional status. Local and systemic sensing of phosphate status, for instance, is not understood, while the regulation via miRNAs is well established (Fig. 7.15).
The knowledge on nitrate perception is most advanced and can therefore serve as an example. The nitrate transporter NRT1/CHL1, already introduced as a protein able to switch affinity in a phosphorylation-dependent manner (Fig. 7.16), functions, in addition, as a nitrate sensor (Ho et al. 2009). Both the primary response to nitrate availability (the up-regulation of nitrate transporter, nitrate reductase and nitrite reductase genes) and the stimulation of lateral root growth in nitrate-rich soil patches (Fig. 7.18) are dependent on NRT1/CHL1 in A. thaliana.
This association is not explained by transporter activity of NRT1/CHL1. Instead, external nitrate regulates the phosphorylation status of the protein and its ability to activate responses to high external nitrate availability. NRT/CHL1 is the first plant example of a so-called transceptor, a transporter protein that in fact functions like a receptor. The nutrient is not a substrate but a ligand with a signalling function. Many similar proteins may be involved in other nutrient-sensing processes.
Date added: 2025-01-27; views: 453;