Nutrient Acquisition Symbioses. Mycorrhizae
For most plants in natural habitats, symbiotic associations with microorganisms are an essential part of their nutrient acquisition. Plants provide reduced carbon to the microorganisms and receive macro- and micronutrients in return. The importance of these symbioses (mycorrhizal associations between plant roots and fungi, and the biological N2 fixation by symbiotic bacteria living within plant tissues) for global biogeochemical cycles cannot be overestimated.
Here, we will describe:
- The mechanisms of nutrient and carbon exchange between the symbiotic partners
- The molecular events leading to establishment of the symbioses, which share so many features that the concept of a “common sym pathway” has emerged.
Mycorrhizae. About 90% of all land plants—most monocots and dicots, and nearly 100% of gymnosperms— engage in a mycorrhizal association with fungi. On the fungal side, tens of thousands of different taxa are involved. Mycorrhizae are present in most terrestrial habitats colonised by plants. Rare exceptions include very dry soils, waterlogged soils or extremely phosphorus-impoverished areas where plants with cluster roots dominate (Fig. 7.19).
Also, most members of a few plant families (e.g. Brassicaceae, Chenopodiaceae) do not form mycorrhizae. Mycorrhizae are evolutionary ancient and most likely were already instrumental when plants first colonised land about 450 million years ago (van der Heijden et al. 2015). This is suggested by fossils of the earliest land plants, which show signs of mycorrhizae.
Two major types are distinguished, which vary in morphology, type of partner and physiology. An endomycorrhiza is characterised by growth of fungal hyphae within the root cortex of the plant host and the formation of structures within cortex cells, while in an ectomycorrhiza the fungal hyphae do not grow into root cortical cells. Ectomycorrhizal fungi sheath the root with a mantle of tightly woven hyphae and grow between cortical cells (Hartig’s net) (Fig. 7.20).
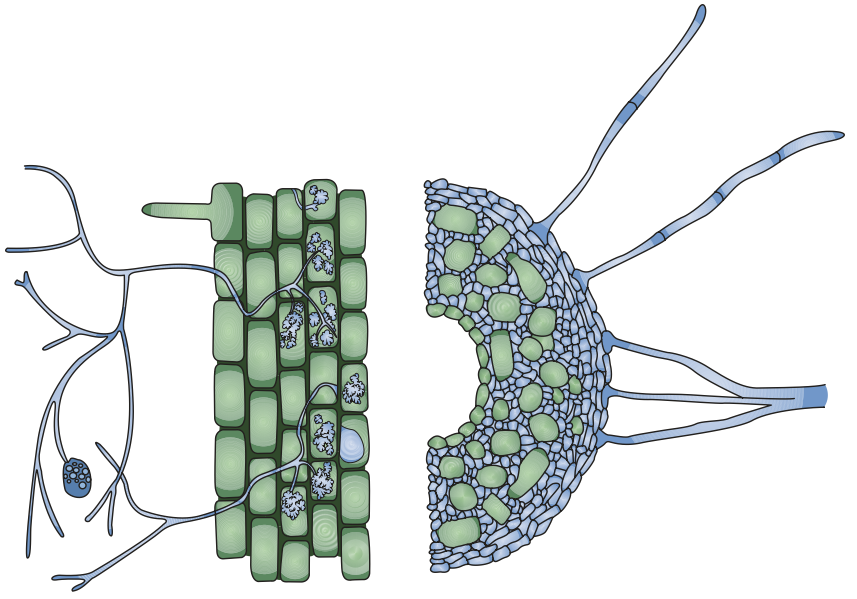
Fig. 7.20. Structural features of an arbuscular mycorrhiza (left) and an ectomycorrhiza (right). (Marschner 2012)
Within the endomycorrhizal symbioses, three types are differentiated further: the arbuscular mycorrhiza, ericoid mycorrhiza and orchid mycorrhiza. By far the most common form (occurring in around 75% of all plant species) and evolutionary the oldest is the arbuscular mycorrhiza. It is also the one that has been studied intensively at the molecular level and will thus be the focus of this section.
A large fraction (up to 80%) of the main limiting macronutrient, phosphorus, is provided to plants by mycorrhizal fungi. Phosphorus is much less mobile in the soil solution than nitrogen or other macronutrients. Therefore, the ability of fungal hyphae to grow into soil micropores that are too small for the diameter of plant roots makes more phosphorus available for uptake. Like plant roots (Fig. 7.5), mycelia actively mobilise phosphorus bound to soil particles—for instance, by the secretion of phosphatases. Zn is another comparatively immobile nutrient that is supplied to the host by the fungal partner. However, this is not well understood. The contribution of the arbuscular mycorrhiza to nitrogen supply for the host is much less pronounced. This is different for ericoid mycorrhizae and ectomy- corrhizae. In addition to nutrients, the fungal symbiont supplies water to the host.
Both the molecular physiology of mycorrhizal transport processes and the events that enable symbiosis formation are being investigated predominantly in the model systems Medicago truncatula and Lotus japonicus. These species are studied in place of A. thaliana which, as a member of the Brassicaceae, does not engage in mycorrhizal symbioses. The added benefit of M. truncatula and L. japonicus as model species is that they are legumes; therefore, biological N2 fixation can be studied as well.
Fungal arbuscules are structures with a large surface within root cortical cells (Fig. 7.21). Thus, the exchange of nutrients and sugars between the fungus and the plant occurs through two membranes that are in close vicinity: the invaginated plant-derived periarbuscular membrane, which is continuous with the plasma membrane of the host cell; and the plasma membrane of the arbuscule.
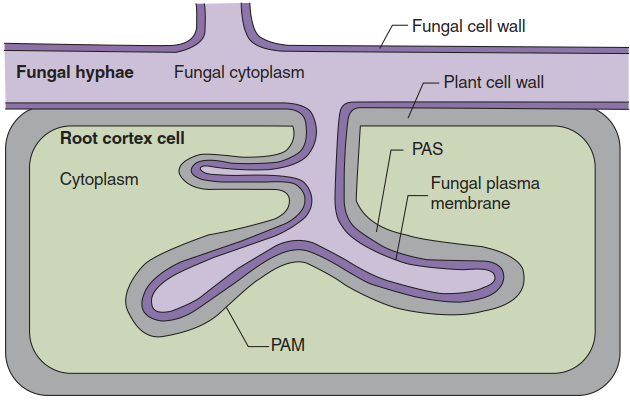
Fig. 7.21. Structure of an arbuscule. An arbuscular mycorrhiza is characterised by arbuscules developing inside root cortex cells. Exchange of sugars and mineral nutrients between the symbiotic partners occurs across the fungal plasma membrane, the periarbuscular space (PAS) consisting of plant and fungal cell wall material, and the plant-derived periarbuscular membrane (PAM), which is continuous with the plant plasma membrane. (Modified from Parniske (2008))
Nutrient transfer from the soil into host cells requires uptake by the fungal mycelium, efflux into the symbiotic interface (the periarbuscular space) and uptake by the plant cell. Transfer of sugars arriving via the phloem from the plant to the fungal partner entails two membrane passages: efflux across the root cell plasma membrane and uptake by the arbuscular membrane.
Experimentally it is extremely challenging to detect and identify the transporters mediating these different steps. Consequently, a molecular dissection of the exchanges between the symbiotic partners is far from achieved. Sequencing of the genomes of plants and mycorrhizal fungi, as well as transcriptomes of mycorrhized and non-mycorrhized plants, is establishing inventories of transporters that are potentially involved (Garcia et al. 2016). The understanding of phosphate transport is most advanced. In host plants, genes encoding specific subgroups of phosphate transporters (the Pht1 family; Sect. 7.3.4) are up-regulated upon fungal colonisation of the root. In turn, other transporters involved in direct phosphate uptake from the soil solution are down-regulated. This indicates that phosphate supply through the symbiosis is prioritised.
How this regulation occurs molecularly is not known. The up-regulated phosphate transporters in colonised root cells take up phosphate from the interface. Energy is provided, as in the case of anion uptake from the soil solution, by the pumping of protons. The proteins involved in the release of phosphate from the arbuscules are unidentified. Uptake of phosphate from the soil solution into the mycelium is mediated by high-affinity transporters similar to the ones expressed by plants (Harrison and van Buuren 1995). An equally fragmentary aspect is the assignment of essential sugar transport steps—that is, efflux of glucose out of the colonised root cell and uptake into the fungal mycelium—to particular proteins.
Even less is known at the molecular level about the possible exchange of reduced carbon, macronutrients and micronutrients between neighbouring plants via mycorrhizal connections. The host specificity—especially of the fungi involved in arbuscular mycorrhizae, the Glomeromycota—is low. Thus, they can be simultaneously associated with individuals of several different plant species. Also, many plant roots are colonised by more than one fungal species. The resulting underground hyphal network may well enable, for example, the support of shaded tree saplings with assimilates supplied by connected adult trees (van der Heijden et al. 2015).
Also, the first direct evidence of carbon trading between trees has recently been found through carbon isotope labelling. A substantial fraction of the carbon isotope label introduced via photosynthesis into one tree was later found in neighbouring trees, suggesting an exchange mediated by ectomycorrhizal connections (Klein et al. 2016). The exact fluxes in such underground networks, however, have not really been determined yet, and the relevant molecular knowledge (e.g. about responsible transporters) is not yet available.
Date added: 2025-01-27; views: 482;