Intracellular Transport and Cellular Aspects of Long-Distance Transport
Nutrient supply to organelles and vacuolar storage require transporters too. Principally the same transporter types responsible for uptake also mediate transport into organelles (Miller et al. 2009). Examples are phosphate transporters in the Pht family. While Pht1 resides in the plasma membrane and takes up Pi, Pht2 is localised to the plastid inner membrane, Pht3 to the mitochondrial inner membrane and Pht4 to the Golgi compartment to mediate Pi transport into these organelles (Lopez-Arredondo et al. 2014). The vacuole is an important storage site for some nutrient ions, including nitrate and phosphate. Vacuolar storage of nitrate is dependent on NRT2 nitrate transporters and nitrate/H+ antiporters of the CLC family.
Not all nutrients are present in the cytosol as hydrated ions that can readily be accepted as substrates for transporters in organellar membranes. Many micronutrients, with Cu as the most extreme example, are too reactive to be available in this form for interaction with proteins and other molecules (Clemens 2001). According to the Irving-Williams series—that is, the observation that the stability of transition metal complexes with organic ligands generally follows the order Mn(II) < Fe(II) < Co(II) < Ni(II) < Cu(II) > Zn(II) (Irving and Williams 1948)—Cu(II) has a higher affinity for organic ligands than any other divalent cation in biology.
This applies to N-, O- and S-ligands—for example, amino acids, organic acids or thiols such as GSH. Therefore, Cu(II) (and Cu(I) after reduction) have to be chelated by designated metallochaperones that deliver Cu ions to various pumps (proteins in the Heavy Metal ATPase (HMA) family) residing in the membranes of cellular compartments (or the plasma membrane for efflux). It has been estimated that in fact not a single hydrated Cu ion is present in the cytosol of a prokaryotic or eukaryotic cell. For Zn(II) the concentration of the so-called labile pool of ions that can readily be transported or bound is in the picomolar range—that is, several orders of magnitude below the total cellular concentration, which is in the micromolar range. Thus, most Zn(II) ions are chelated either by proteins or by low molecular weight ligands (Sect. 7.5.1). Estimates for the other micronutrients including Fe are less precise (Fig. 7.17).
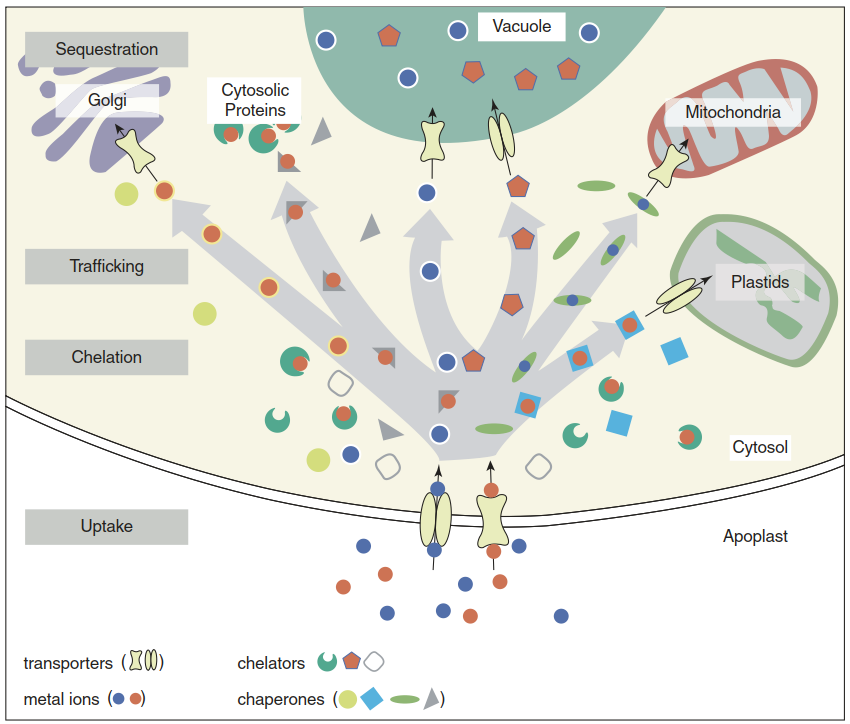
Fig. 7.17 Cellular metal homeostasis. Following uptake through plasma membrane-localised transporters, the ions of Cu and Zn in particular are bound by chelators and chaperones. Chelators buffer cytosolic metal concentrations and chaperones are involved in trafficking metals to target sites—that is, organelles and proteins requiring the metal in question. In particular, Cu chaperones directly interact with metal pumps that transport the metals into organelles
Following symplastic passage from sites of uptake in the root cortex (Fig. 7.13), nutrients are loaded for long-distance transport into the xylem. The xylem is apoplastic. Thus, efflux across the plasma membrane of the xylem parenchyma or pericycle cells is necessary. Energetically the situation is reversed in comparison with uptake. Anions can be loaded via facilitated diffusion; cations may require active transport. Many transporters involved in this loading process are known in the model systems A. thaliana and rice. Loss of function leads to reduced xylem sap concentrations of the nutrient in question. This results in lower shoot concentrations.
In A. thaliana, nitrate, which can (in contrast to ammonium) be transported over long distances, is loaded into the xylem by the low-affinity transporter NRT1.5. The cytosolic nitrate concentration is high enough for this type of transport. NRT1.5 shows the typical localisation of expression in pericycle cells adjacent to the protoxylem to fulfil its loading function (Fig. 7.13). Similarly, borate is exported into the xylem by transporters known from A. thaliana and rice (BOR1). K+ ions reach the xylem through outward-rectifying Shaker-type channels—SKOR in A. thaliana. Its opening probability decreases with rising external K+ concentrations. This ensures that only efflux is mediated (Cherel et al. 2014). Zn is an example of a nutrient that is actively pumped into the xylem. The proteins responsible are HMAs. In A. thaliana, HMA2 and HMA4 load Zn into the xylem.
Date added: 2025-01-27; views: 439;