The Biological Evolution of Vultures. Convergent evolution; What is it?
Introduction. This chapter examines the history of both groups of vultures, from the earliest, disputed forms, to the more recognizable species that spawned the Old and New World vultures. The origins of the New World vultures are more disputed than those of the Old World vultures. The New World vultures have been classified within the Acipitridae, Ciconidae, and also independently, in their own Order. The Old World vultures, by contrast, are seen as evolving within the Accipitiformes.
At the beginning of vulture evolution, five main issues stand out. First, the earliest 'vultures' are not necessarily recognized as true 'vultures'. Here, a vulture is defined as a largely carrion eating bird, with feet not as clearly designed for killing as those of hawks and eagles, with bare or semi-bare heads, broad wings for soaring flight and bills designed for either pulling, tearing or picking. Second, what is termed Old World or New World vulture is less definable by location, as Cathartid ancestral groups have been discovered in the Old World and the Accipitrid vulture ancestors are fossilized in the Americas. As noted by Zhang et al. (2012b: 7) 'birds fly, and their ancestral distributions may not correlate with their modern distributions.'
Third, the classification of fossils, based on skeletal form compared with modern forms, does not conclusively determine the diet of such birds. For example, an extinct counterpart of the Black vulture could be a vegetarian, and as such its 'vulture' status would be disputed. Fourth, much of the evidence is circumstantial; merely because one bird form was dated to a period before another similar form, does not conclusively prove that the latter evolved from the former. Fifth, vultures may not necessarily have reached their final form; the new ecosystems they inhabit, with less food and human modified landscapes may encourage further convergence or divergence.
With these ideas in mind, this chapter first looks at the prehistoric scavengers that are considered ancestral to modern vultures. It follows up with a detailed consideration of the New World and Old World vultures.
Convergent evolution; What is it? New and Old World vultures are generally seen as an example of convergent evolution. Critical examination of the biochemistry, morphology and anatomy of Old and New World vultures shows that the two groups 'represent a phylogenetically inhomogenous, i.e., polyphyletic group, whose shared characters are based on convergence' (Wink 1995; as also noted by Mundy et al. 1992; Sibley and Ahlquist 1990), which some have called a 'textbook example of convergent evolution' (Del Hoyo et al. 1994).
Both groups eat carrion, but Old World vultures are in the eagle and hawk family (Accipitridae) and use mainly eyesight for discovering food; the New World vultures are of obscure ancestry, and some (Genus Cathartes) use the sense of smell as well as sight in hunting. Birds of both families are amongst the largest flying birds in the world, have unfeathered heads and necks, search for food by soaring, the larger species live in mountains, all circle over a sighted carrion, nest in tall trees and high cliffs, and may roost in flocks in trees. Other shared characteristics are long intestines, strong stomach acids to digest rotten meat and bones and kill microorganisms in carrion, feet for movement on the ground, and lack of the raptors' pronounced sexual dimorphism (Newton 1990; Brown and Amadon 1968; Mundy et al. 1992; Glutz von Blotzheim et al. 1971; Cramp and Simmons 1980; del Hoyo et al. 1994).
Such similarities are apparently possible, if species from different Orders and Families slowly adapt to similar opportunities and through time, resemble each other in both appearance and behavior. For example, all avian carrion eaters will need soaring flight, as carrion exists in widely spaced areas, and soaring birds outcompete terrestrial bird or mammal scavengers. They may need bare heads and necks (even the Marabou Stork (Leptoptilos crumenifer, Lesson 1831), a habitual carrion-eater, has a bare head to avoid bloodied feathers); they may be large, as speed is less important than the ability to rip open carcasses, and to store large quantities of food in their crops against lean seasons; and they may not need the strong feet of predatory eagles and hawks, for terrestrial locomotion. Any Families, such as hornbills, gulls, storks or raptors that gradually adapt to this lifestyle, possibly due to the abundance of carrion (such as during the Age of Mammals) and the relative ease of carrion consumption compared with catching live prey, might gradually acquire similar appearance and habits.
All vultures were once thought to have evolved 'only once among extant diurnal birds of prey' (Sielbold and Helbig 1995: 163). But in the late 19th century, scientists queried the status of the Cathartidae as vultures (Garrod 1873; Sibley and Ahlquist 1990). Rea (1983: 26) argues that 'from the very beginning the relationship of the cathartid vultures to the Falconiformes has been controversial.' Further, 'so far in the fossil records of Cathartids, Accipitrids and Falconids, there is no time when Cathartid vultures were more raptorial or the raptors less raptorial, there is no fossil candidate for a common ancestor' (ibid. 34). Vultures were initially hypothesised to be a monophyletic group, but with the increasing awareness of distinctions between New and Old World vultures, vultures were increasingly seen as polyphyletic. In this sense, a monophyletic group is a taxon (group of organisms) forming a clade, i.e., consisting of one ancestral species and all its descendants. A polyphyletic group has convergent features not inherited from one common ancestor, but from several such species.
Based on studies using anatomy, morphology and biochemistry, there is evidence that vultures must represent a phylogenetically diverse group (i.e., they share different ancestors; a polyphyletic group) and the shared characteristics are due to convergence (Mundy et al. 1992; Sibley and Ahlquist 1990). Del Hoyo et al. (1994) even called them a 'textbook example of convergent evolution'. A study using biochemical nucleotide sequences of the mitochondrial cytochrome b gene (which holds information on the biological relations among animals) found evidence that the Cathartidae (New World Vulture family) was not closely related to the other raptors in Acipitridae (including the Old World Vultures). 'Since also the fossil record provides no evidence that the different families within the Falconiformes had a common ancestor, 'raptors' should be a result of an evolutionary convergence between bird groups of polyphyletic origin'; the similarities 'found in morphology are indeed based on convergence and not on close genetic relatedness' (Wink 1995: 880). The evidence for convergence, found by morphological (the study of the appearance and shape of the skeleton and body), karyological (the study of chromosomes) and DNA-based studies show vultures are in at least three monophyletic clades (this means that each group possibly has a different common ancestor) showing polyphyly (multiple ancestors) (see also Sibley and Ahlquist 1990; Mundy et al. 1992; Del Hoyo et al. 1994).
Convergent evolution is the independent evolution of similar or identical features in species of different Genera, Families or Orders. In convergent evolution, similar structures with similar form and/or function that were non-existent in the ancestral groups, may develop in the descendant groups. These similarities are termed analogous structures, which differ from homologous structures (which may share a common origin but have different functions). Convergent evolution is the opposite of divergent evolution, in which similar, related species develop different traits. The most similar phenomenon to convergent evolution is parallel evolution. The latter occurs when two independent but similar species change in similar ways. The difference lies with the ancestors; if the ancestors were similar then the evolution is parallel, if they were different, then the evolution is convergent.
Problems emerge when the ancestral forms are not definitively known; in this case parallel and convergent evolution are less distinguishable. Further complications emerge when different structures perform a similar function. It is also possible that where lineages do not evolve together and then do so, the result may be convergent evolution at some later point. For example, if all birds share a common ancestor, then evolved into different species in different places (for example, into raptors and storks), later convergent evolution could create the two groups of vultures, but there may have been parallel evolution at some earlier point in time.
It must be noted that the definitions of traits may also determine whether a form of change may be considered divergent, parallel or convergent. For example, if we consider only the heads of vultures, there is convergent evolution of bald heads. Likewise, the broad wings for soaring flight indicate convergent evolution. As also shown in Fig. 2.9, there are peckers and tearers among both the New and Old World vultures, with few differences between the Egyptian and Hooded vultures on one side and the Cathartes and Black vultures on the other. The condors and King vulture may be seen as tearers, although their bills are proportionately slightly smaller than the giant bills of the Lappet-faced, Cinereous and Red-headed vultures. However, other traits show divergence, such as the weaker feet of the New World vultures (see Fig. 4.1 for a comparison of vulture, eagle and stork feet), their habits of defecating on their legs and feet, their perforate nostrils and in the Cathartes vultures, the sense of smell.

Fig. 4.1. The Feet of Vultures, Eagles and Storks
The pronounced brow ridges above the eyes of the Old World vultures are more similar to raptors like eagles, while the small brow ridges of the New World vultures (the eyes appear to pop out of the skull) are more similar to the storks (Fig. 4.2). These would count as divergence if they shared a common ancestor with the Old World vultures, as the latter do not have these traits. Taking all the traits of both groups together, the result is termed convergent because it is believed that there are distinct ancestral groups.
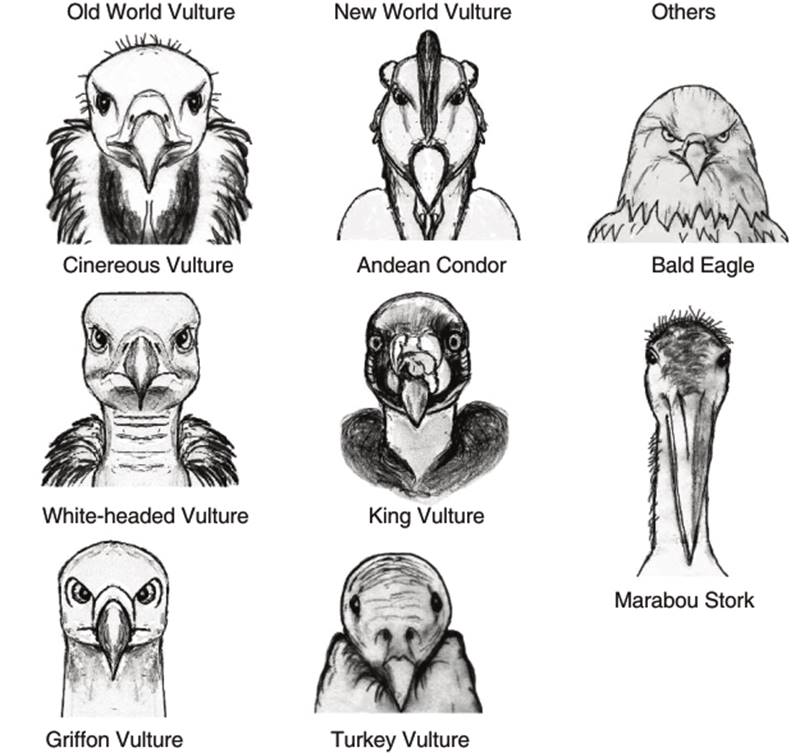
Note the larger, eagle-like brow ridge above the eyes in the Old World Vultures compared with the stork-like head and face of the New World vultures
Fig. 4.2. The Heads of Vultures, Eagles and Storks
Rea (1983) notes that the New World vultures differ from the Old World vultures and other raptors in the following: mycology of thigh and breast; pteryography; absence of patagium connecting the secondary flight feathers; arrangement of wing coverts; lack of syrinx and intrinsic syringeal muscles; in the configuration of the laryngeal apparatus; the shape of wing claw; relative sizes of liver lobes; thermoregulation and nest defence; and chromosomes, hematology, egg white protein and physical structure of eggshells. This author (p. 45) states 'there is nothing in the fossil history to unite the groups or to suggest that one might be derived from the other.' Furthermore, the Catheridae must be placed with Ciconiiformes, as 'the most inescapable conclusion from an examination of the Catheridae is that they have virtually no trait, whether anatomical, physiological, cytological or behavioral, that suggests that they are related to the Falconiformes.'
An important issue in the study of vulture biology concerns the lack of sexual dimorphism in both the New and Old World vultures. In raptors, the female is much larger than the male, but in both condors the male is slightly larger, and in many other vultures the distinction is minimal (although size may favor the male in New World vultures, and the female in the Old World vultures). This makes it difficult in some cases to assess the population structure and ecological behavior (Fry 1983). Methods used to distinguish between male and female birds include: direct gonadal (sex organs, testes and ovaries) examination by laparotomy (incision) or laparoscopy (incision with a camera); analysis of blood or urinary steroids; karyotype analysis through identification of sex chromosomes; and behavioral differences between males and females. Some of these methods are more easily used than others. Urinary steroids can be measured from fecal samples, but the other methods require physical handling of the bird to obtain blood samples, and judging sex by behavior is unreliable (Fry 1983).
Date added: 2025-04-29; views: 472;