The Paradox of Biological Individuality: From Genets to Colonies in Evolutionary Ecology
The individual organism figures prominently in this book. As discussed in the Preface, this is largely because natural selection acts primarily at the level of the individual. But what is an individual? The word is ambiguous because it may be used by ecologists in at least three contexts. These are not mutually exclusive and are considered below (Fig. 1.3). In passing, we should note that the related matter of what constitutes ‘individuality’ is not explored in this book, other than briefly with respect to size increase in the volvocine algae in Chap. 4.
The evolutionary point(s) at which a mass of cells becomes sufficiently differentiated, organized, and interdependent to be declared ‘an individual’ is contentious and at the extreme becomes a metaphysical issue. Definitions are subject to various debatable criteria, such as whether or not germ-soma differentiation is a necessary condition. Individuality is considered elsewhere (e.g., Buss 1987; Maynard Smith and Szathmary 1995; Michod 2006, 2007; Herron and Michod 2008; Folse and Roughgarden 2010; Szathmary 2015). Several of the ideas may lend themselves relatively well to animals but not to plants or microbes, for example because of the totipotency of their cells.
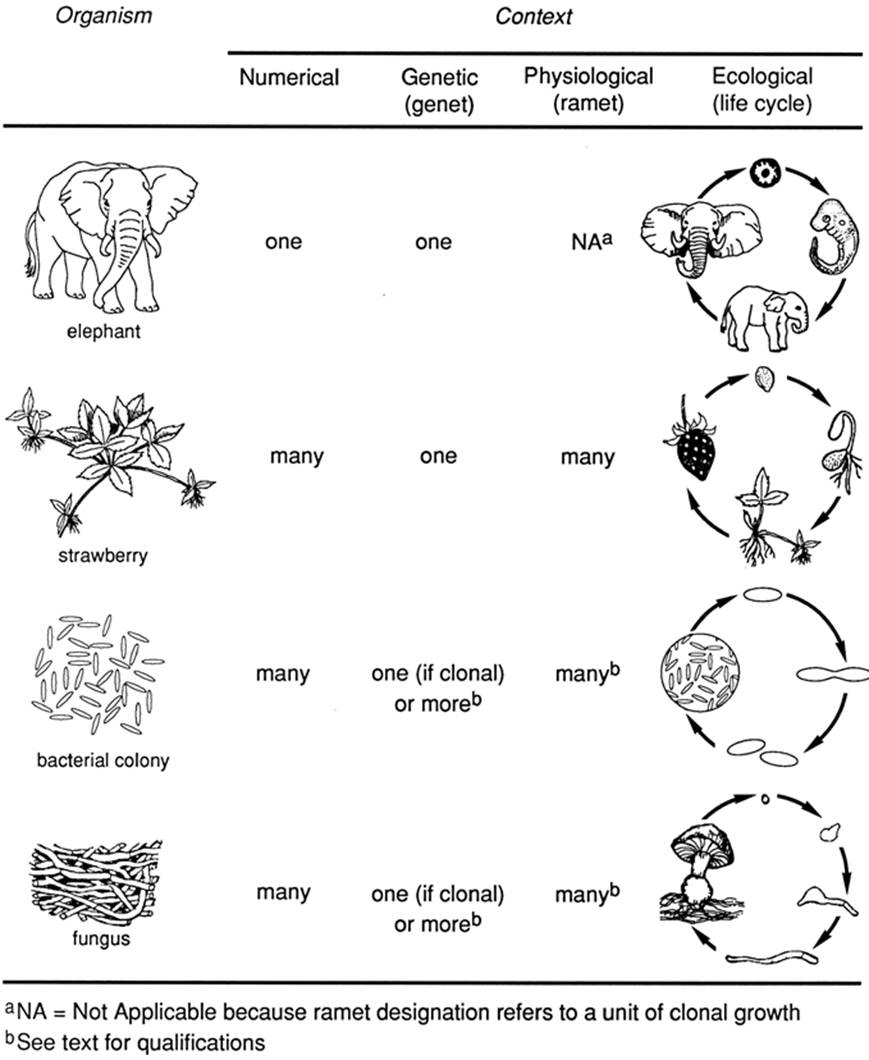
Fig. 1.3. Relationships among different concepts of an individual for various kinds of organisms
With respect to the term ‘individual’, first, it is often used operationally in a numerical or quantitative sense to mean a representative of a particular population or species, something that can be counted. For some organisms this is intuitively clear: we can all visualize one rabbit or one maple tree. Individuals by this definition are supposedly discrete and functionally independent units. For colonial or clonal organisms such as bacteria, fungi, various algae, bryozoans, and coelenterates, however, it is not at all clear what level of cellular aggregation fulfills these criteria (e.g., Larwood and Rosen 1979).
For example, the reason the term CFU (colony-forming unit) is used in bacteriology and to some extent in mycology is that in practice it is generally unknown whether growth (as on solid medium in a petri dish) is produced from one cell or a cluster of cells, or whether all potentially living cells seen microscopically in a sample will multiply in the appropriate medium.
Second, the term ‘individual’ may be used in a genetic sense to mean the entire unit, abbreviated genet, resulting from growth of a zygote (Fig. 1.3; this idea is developed in Chap. 5; see also Harper 1977, p. 26; Anderson and Kohn 1998). This usage is equivalent to the numerical definition when applied to unitary organisms such as most mobile animals (Chap. 5), that is, those in which the zygote develops into a determinate body repeated only when a new life cycle starts. A single deer is both a numerical individual and also technically a genet (though rarely referred to as such).
The genet concept is meaningful in evolutionary terms because it focuses attention on the genotype through time. It came into vogue because, for modular organisms such as corals and those plants with clonal (asexual) growth, the number of discrete, countable 'individuals' is not the same as the number of genets (Kays and Harper 1974). The term ramet is given to each of these countable units (Chap. 5; see also Harper 1977, p. 24). Each is a member of a genet but is actually or potentially capable of independent existence as a separate or physiological individual (Fig. 1.3). The case for carrying over the terminology and concept to microorganisms is argued in Chap. 5.
Thus, it follows that ramet only has meaning within the context of organisms wherein the genet is composed of multiple, independent parts and therefore, by definition, does not apply to unitary organisms. As such, ramet appears under its own heading in Fig. 1.3 with the relevant entries for each organism example. An entire hillside of bracken fern or field of dandelions may constitute a single genetic individual (one genet or clone), being nothing more than the multiple phenotypic representation (many ramets) of one successful genotype (Harper 1977, p. 27; Janzen 1977b). Of course in practical terms, if several genets intermingle, as is often the case, they may well appear superficially identical, and usually there is no easy way to differentiate visually among them. As taken up at length in Chap. 2 and subsequently, members of a clone diverge genetically more or less quickly over time. The real issue is the rate at which this occurs and whether the genetic changes are significant functionally (evo- lutionarily). For example, depending on the ontogenetic program of the organism, mutations that occur in somatic cells may enter the germ line (Buss 1985), which, from an evolutionary standpoint is the critical issue. How the genetic individual relates to the numerical entity is an ongoing and fascinating problem in ecology and evolutionary biology. It is a recurrent theme in this book, a foretaste of which is the paper by Pedersen and Tuomi (1995), who consider the levels of organization in modular and clonal organisms.
In summarizing the above quandary, Allen and Hoekstra (1992) say amusingly and insightfully, in part ... (p. 160) “the archetypal organism is human, and other organisms variously represent departures from ourselves, roughly in the order: cuddly and childlike; warm but big; scaly and cold; immobile; microscopic. The further away from being human is the organism in question, the less do the more formal attributes of organisms apply.”
Third, in an ecological sense, the individual—though usually thought of in the adult form rather than as a juvenile or zygote—can be taken implicitly to be the organism through its entire life cycle (Fig. 1.3). Within the life cycle context, an organism is a whole and this entity can be visualized in its various genetical, physiological, morphological, or developmental states, from birth to death. For unitary organisms, there is a direct and clear correspondence among these three definitions—life cycle, numerical, and genetic—from birth to death or from zygote to zygote. Genetically different individuals (i.e., new genets) are produced at one or more specific points in the life cycle. Germ cells in the parental genet undergo meiosis to produce gametes, sexual reproduction occurs, and the new genets emerge into the population as developing zygotes.
The parent typically continues to live for some time and consequently generations overlap. This contrasts sharply with the case for some modular organisms wherein the genet undergoes fragmentation, such as in the free-floating aquatic plants (Lemna spp. [duckweeds], Azolla spp. and Salvinia spp. [water ferns]), and the corals. Modules may be sloughed and are free to move about passively. Kariba weed (Salvinia molesta), which propagates clonally, has come to occupy vast areas and may weigh millions of tons (Barrett 1989). Thus, it considerably exceeds the mass of the blue whale (the largest extant unitary organism on earth), but may be challenged for this record by the fungi (Smith et al. 1992)! Of course the numerical as well as genetic individual in the case of microbes tends not to be discrete in either time or space; indeed the microbial clone is often widely and occasionally globally distributed.
In summary, there is no simple, unambiguous concept of an individual that can be used across the spectrum of life as a benchmark in the comparative ecology of micro- and macroorganisms. Comparisons can be made conceptually to a greater or lesser extent by use of any of the three definitions depending on context, but in a strict sense not functionally. Definition one, the numerical usage, is rejected here as a useful common denominator because it means little in evolutionary terms and operationally in microbiology is quite contentious, notwithstanding the application of molecular methods, which in the past two decades have largely overcome limitations such as the ‘viable but uncultivable’ problem discussed in later chapters (Brock 1971; Colwell and Grimes 2000; Walker and Pace 2007). Thus, to arbitrarily choose individual bacterial cells, fungal colonies, and slime mold amoebae, and attempt to compare them with individual trees, insects, and birds would be futile.
To the extent that the genetic individual, definition two, implies simply that a genet is the product of a zygote, it can be applied widely, in many cases literally and in others conceptually where a true zygote is not strictly produced. But it should not be mistaken to imply genetic homogeneity, i.e. absence of sub-organismal variation. A key difficulty posed by microbes is that an individual cell or clone can change genetically in a manner different from the conventional process of reproduction with meiosis, so it may be impossible to know when the genetic individual starts to exist or when it ends. As seen above, for unitary organisms it begins at the time of genetic fusion, in other words, when genetic recombination occurs at zygote formation and the contribution from each parental genome is diluted by exactly one half.
Moreover, the point in the life cycle when karyogamy can occur is fairly standard and usually easily recognized and associated with particular terminology such as reproductive phase; sexual maturity; seed production. This is not the case for bacteria and fungi, which display indeterminate, highly plastic development and experience novel genetic events. A bacterial cell may divide repeatedly, yielding descendants that may be relatively sequestered as an initially genetically homogeneous, clonal microcolony, albeit one that changes progressively by mutation (Milkman and McKane 1995). Moreover, horizontal (lateral) gene transfer may occur through various atypical sexual processes (7-Chap. 2). How many genes have to mutate, be acquired, or lost in a haploid organism for our conceptual genet to effectively cease to exist?
A roughly analogous, ambiguous phenomenon occurs in the fungi that have a dikaryotic (n + n) phase or exhibit parasexuality (Chaps. 2 and 6). For instance, Puccinia graminis tritici, the parasite causing stem rust of wheat, has five spore stages with associated mycelial growth involving both barberries and wheat as the alternate host. The life cycle includes haploid, dikaryotic, and diploid phases. Shortly after meiosis, the fungus becomes partially or fully dikaryotic by various mechanisms (Chap. 6), but karyogamy is delayed and true diploidy is a transient stage. The dikaryotic phase, technically haploid but functionally diploid, is of variable duration. Furthermore, in absence of the barberry as an alternate host but in the presence of a favorable environment, the fungus may cycle indefinitely in the dikaryotic condition. What, then, is the genet for the wheat rust fungus? Whatever the debatable answer, the genet corresponds neither to a single numerical entity, nor is the ploidy static throughout the full cycle. This is a much more complicated situation than occurs with, say, a clonal buttercup or an aclonal maple tree, and may be most closely approximated by plants that have an extended gametophyte stage such as the fern.
The genet concept could be brought into line with what intuitively seems to physically represent the organism by allowing for a certain amount of genetic flux after recombination in the zygote, or at the start of the life cycle in the case of haploid and asexual organisms. This would recognize that, while the genetic basis for the individual is established by genetic recombination in some form at the outset, the genotype could be molded by additional genetic events that might periodically occur during the life cycle. As noted above, in bacteria this can take the form of mutation or HGT (horizontal gene transfer, Chap. 2). For plants and benthic invertebrates it can take the form of somatic mutations, some of which may enter the germ line. However, the full evolutionary consequences remain to be seen.
Definition three, the individual depicted as an organism passing in more or less organized fashion through stages in its life cycle, provides a reasonable, universal basis for ecological comparisons. Unless noted otherwise, this is the frame of reference that is used throughout this text. The scheme is applicable to all of the various phases of diverse organisms, and accommodates changes in ploidy, morphology (phenotype) including size, and variation in the time and extent of genetic change. Bonner (l965) has interpreted the life cycle as proceeding sequentially from a state of minimum size (generally at the point of fertilization) to maximum size (generally the point of reproductive maturity). The transition involves growth over a relatively long period to adulthood, followed by return to minimal size over a relatively short period, accomplished by separation of buds or gametes from the adult. For single-celled organisms, the cell cycle is the life cycle of the physiological but not of the genetic individual. These matters are taken up at length in Chap. 6 for which the stage is set in the next chapter.
Date added: 2025-06-15; views: 360;