Origins, Geobiochemistry, and Phylogenetic Benchmarks
There is complementary, reasonably strong evidence from geology and astronomy that Earth originated about 4.5 Gya (Gya = Giga or billion years ago) from a collision of planetesimals (Nisbet and Sleep 2001; Knoll 2015). Following a period of cooling, a crust was formed and temperatures allowed the existence of liquid water and other geochemical conditions conducive for the formation of the organic precursors of life. Life probably emerged sometime between the end of the last accretion impacts from meteorites (~4 Gya) and the formation of the earliest chemical fossils, which date to about 3.8 Gya (Barton et al. 2007). The broad sweep of early evolution of life on Earth is summarized in Fig. 4.2. It should be kept in mind that all such reconstructions are to varying degrees contentious and whether based on combined or separate fossil and molecular approaches are subject to various uncertainties, caveats or constraints, assumptions, and degrees of inference.
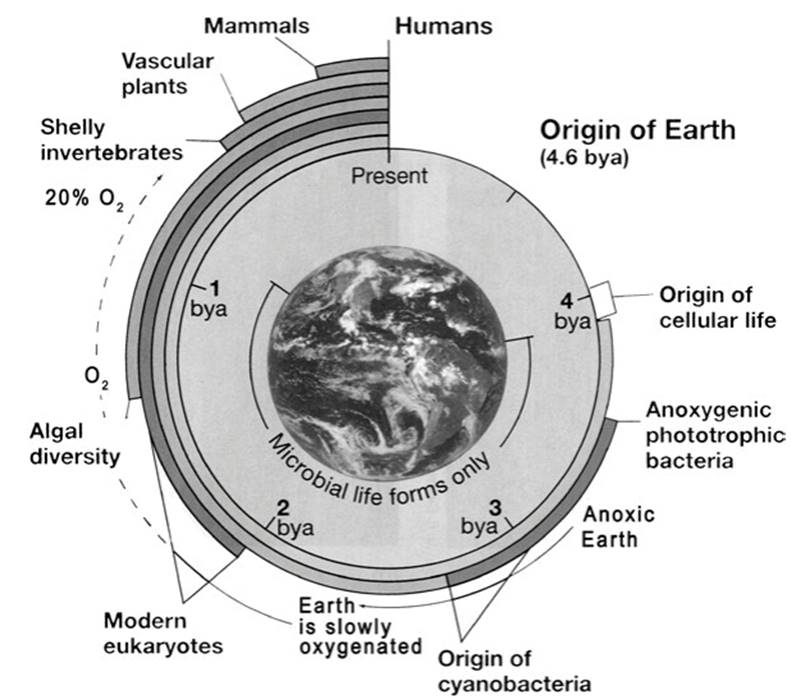
Fig. 4.2. Overview of the history of life from the origin of Earth 4.5-4.6 billion years ago (bya or Gya) to the present. Cellular life probably arose ca. 3.8-4.0 bya and for half or more of the history of life on Earth was exclusively microbial. Very early life forms were anoxygenic chemotrophs and phototrophs followed by oxygenic (oxygen-generating), phototrophic cyanobacteria, which set the stage for evolution of increasingly complex, multicellular eukaryotes. For synopsis, see Hedges (2002). Reproduced from: Madigan, Michael T.; Martinko, John M.; Stahl, David A.; Clark, David P. Brock Biology of Microorganisms, 13th Ed. ©2012. Printed and electronically reproduced by permission of Pearson Education, Inc. New York, New York. Image of Earth from space produced by M. Jentoft-Nilsen, F. Hasler, D. Chesters (NASA/Goddard) and T. Nielsen (Univ. Hawaii)
A cornerstone of the current scientific paradigm for the early development of life is the so-called 'Universal Ancestor' (UA), which is a phylogenetic term for origin of the ‘tree of life’ and the nebulous entity(ies) from which all known life presumptively evolved (Woese 1998a). A common evolutionary origin is the most direct (parsimonious) inference from the fact that living creatures exhibit universal homologies, most notably a standard biochemical imprint of informational and building block molecules and cellular attributes summarized in Chap. 3. Arguably, the UA was not a singular entity as visualized by Darwin, but may have been a fluid community of diverse, freely intercommunicating primordial cells variously termed precells, protocells, or progenotes.
Although the word 'progenote' has come to represent different things, as originally articulated by Woese in several papers (e.g., 1987, 1998a, b) it was, as distinct from “genote”, a cellular construct that had an incomplete linkage between its genotype and phenotype and other features such as a short-stranded RNA as the information molecule; few disjointed genes; no cell wall; simple metabolism; and free cross-feeding with the environment and other protocells. Interestingly, if correct, this interpretation implies that contemporary horizontal gene transfer in prokaryotes and eukaryotes (Chap. 2) may be a vestige of the profligate rudimentary horizontal transfer of the primordial world.
There is no unambiguous fossil record for the cryptic progenote and placement of the era is by extrapolation. The lower boundary is set from the estimated age of ancient prebiotic assemblages obtained by geologic biosignatures in rock dated radiometrically; the upper boundary is marked by the first occurrence of putative stromatolites indicative of microbial (mainly cyanobacterial) activity, some arguably as old as ~3.4-3.7 Gya (Tice and Lowe 2004; Schopf et al. 2007; Nutman et al. 2016). Nisbet and Sleep (2001, p. 1086) define stromatolites as “organosedimentary structures produced by microbial trapping, binding, and precipitation, generally but not always photosynthetic”. The reliability of many such early traces is vigorously contested (Brasier et al. 2006).
Indeed, as alluded to above, all models for the origin of life are controversial, but in most of them the UA preceded the first simple, unicellular prokaryotes. This means that it would have been older than ~3.5 Gya. As noted at the outset, the earliest chemical fossils date to about 3.8 Gya in the late Archaean Eon (3.8-2.5 Gya) with ostensible biosignatures and microfossils variously attributed to hypothesized anoxygenic phototrophs or iron/S oxidizers by the mid-Archaean.
Geochemical markers indicative of oxidized rock imply that atmospheric oxygen levels, attributable largely to the photosynthetic activity of cyanobacteria and sufficient to support aerobic respiration, were significant by 2.8-2.2 Gya (Nisbet and Sleep 2001; Bekker et al. 2004). In other words, for about the first billion years Earth was anoxic. Oxygen availability was a critical event for several reasons, in major part because the absence of complex eukaryotes has been attributed to limited oxygen (King 2004). Presumptive multicellular, colonial macrofossils up to 12-cm in size and ascribed as living aerobically and possibly in mat-like arrays have been reported from 2.1 Gya (El Albani et al. 2010). This report is debatable and it is unclear whether the structures, if representing macroscopic life, are prokaryotic or eukaryotic. Unicellular eukaryotes (protists) arguably arose more or less concurrently with mitochondria, likely by ~1.5 Gya and possibly much earlier (Dyall et al. 2004). Pace (2009) infers based on molecular phylogenies that the eukaryote nuclear line of descent is as old as the archaeal, i.e., that the two forms have been present since the beginning of recognized life.
What such an incipient eukaryote may have looked like is unclear. Fossils interpreted as presumptive eukaryotes are dated at 1.45 Gya (Javaux et al. 2001) and an authoritative benchmark for a multicellular red alga (implying the presence of both mitochondria and plastids) was recovered from sedimentary rock in the Canadian arctic dated at 0.75-1.25 Gya (Butterfield et al. 1990; Butterfield 2009). Molecular phylogenies now suggest that the green algae date to at least 1.5 Gya (Yoon et al. 2004). Thus the fossil and molecular evidence point to a major burst of multicellular eukaryotes dated about 1 Gya (King 2004; Grosberg and Strathmann 2007), including the multicellular ancestor of modern animals at about 0.6 Gya. Thus, remarkably, for half or more of the history of all life on Earth the biota was exclusively microorganisms, and overwhelmingly if not exclusively, prokaryotes.
How did the earliest life forms provide for themselves? Among the innumerable possibilities, there are three classical hypotheses (not mutually exclusive) for their functional or trophic nature. Because of the complexity of chlorophyll or similar pigments as well as of the associated energy transduction pathways, all three models visualize the incipient forms as being anaerobic chemotrophs rather than phototrophs. Beyond that, they are postulated as being either: (i) organoheterotrophs (“prebiotic or organic soup theory”; Bada 2004); (ii) lithoautotroph producers, possibly hyperthermophiles (“metabolist theory”; Wachtershauser 2006); or (iii) lithoheterotroph organic conservers (strictly neither producers nor conservers; Ferry and House 2006; for terminology see Chap. 3). Whatever their exact nature, all would have required then, as now, chemical substrates both for assimilation into monomers and polymers of cell biomass, as well as for dissimilation for energy generation.
This, in turn, depends on linked redox reactions, i.e., nonequilibrium electron transfers from donors to acceptors. It is logical that because of the complexity of needed incipient biogeochemical pathways and the expense of autotrophic organosynthe- sis, the earliest cells probably acquired their building blocks from geochemical or extraterrestrial sources. The probable route for energy generation was via simple inorganic electron donors such as cyanide, CO, H2, and FeS arising from cold seeps or t hermal vents (for examples, see Amend and Shock 2001; Martin et al. 2008; Knoll 2015). Regardless of the specific scenario, it is evident that emerging catalytic (enzymatic) control of chemical disequilibrium thermodynamic forces was the bridge between chemical and biological evolution. This critical step is probably the most fundamental common denominator in the origin of life.
Date added: 2025-06-15; views: 315;