The Evolution of Life Cycles: From Prokaryotic Simplicity to Eukaryotic Complexity
Historically speaking, the original and simplest life cycles probably arose in unicellular organisms concomitantly with cellular division of labor and the onset of sequential events at the molecular and physiological level. A contemporary analogue would be the cell cycle in undifferentiated, unicellular prokaryotes. That cycle begins with a single cell and ends with two separate, superficially identical, daughter cells (in fact they are not strictly identical, as will be developed later). It should be emphasized in passing that the cell cycle is distinct from the life cycle of the prokaryotic clone, which continues indefinitely (more on this later).
With respect to prokaryotic life cycles and unlike the case with eukaryotes, reproduction (fission) in bacteria and gene exchange are different processes as was reviewed previously. The cycle is typically haploid, since most (not all; see Chap. 2) prokaryotes contain only a single chromosome and therefore a single copy of each gene. Notwithstanding little conspicuous morphological change prior to the actual divisional event, underpinning the process is a regulated temporal pattern in gene expression, biosynthesis, etc. At a slightly more complex level, discrete morphological changes occur in the cell cycle of bacteria that form spores as a resistant state (e.g., Bacillus), or that alternate between a sessile, feeding stage and a motile, reproductive state of two or more cells. This latter group includes, for example, Caulobacter and relatives (Brun and Shimkets 2000; Laub et al. 2000) (Fig. 6.1).
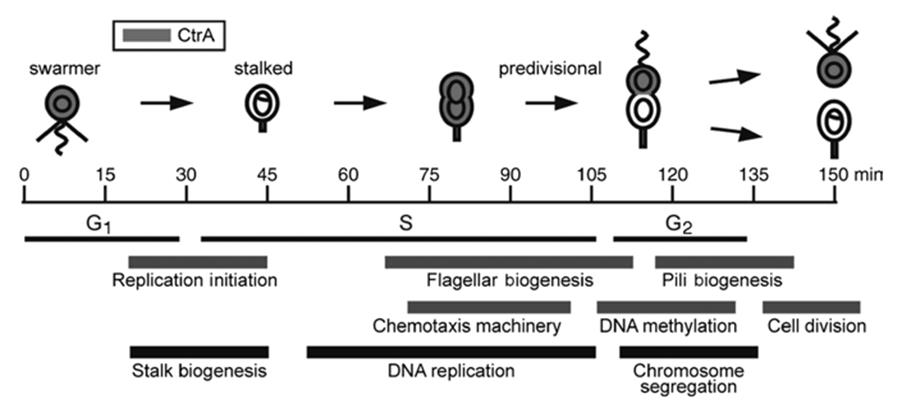
Fig. 6.1. A simple dimorphic life cycle represented by the aquatic bacterium Caulobacter crescentus. Morphogenesis, cell cycle stages (G1, S, G2), and certain associated biochemical activities are shown as a function of time. (The shaded cells contain a response regulator, CtrA.) During each cell cycle a flagellated, swimming cell (swarmer) and an ultimately stalked, sessile cell are produced. Following a motile phase, the swarmer sheds its flagellum and an adhesive holdfast is produced cementing the cell to the substratum, following which a stalk is synthesized at the holdfast pole. Interestingly, through its role in cell elongation the stalk enhances nutrient uptake, advantageous in oligotrophic waters (see Wagner and Brun 2007). From Laub et al. (2000); reproduced from Science by permission of The American Association for the Advancement of Science, ©2000
The molecular genetic underpinning of this Caulobacter life cycle and the morphological transitions, particularly with respect to regulatory proteins, are known to some extent and are probably quite analogous in both prokaryotes and multicellular eukaryotes (Jiang et al. 2014). Other prokaryotes may exhibit multiple alternative states (myxobacteria discussed in 7Chap. 4) or differentiate specialized cell types (e.g., akinetes and heterocysts in the filamentous cyanobacteria) (Dworkin 1985).
Thus, ‘development’ in the bacterial cycle consists largely of significant transitions in function and, in some species, the morphological changes between alternative cell types in response to environmental signals. This sequencing and the diversity of prokaryotic life cycles are discussed by Dworkin (1985, 2006) and Brun and Shimkets (2000). These seemingly simple changes are the prokaryotic counterpart to morphological complexity and developmental staging in eukaryotes. Remarkably, however complex the life cycle ultimately becomes in multicellular macroorganisms, it characteristically if not universally incorporates, sooner or later, at least one single-cell stage (typically the spore and/or zygote; apparent exceptions to this principle are discussed by Grosberg and Strathmann 2007). The significance of passage through a single-celled stage is discussed in a later section, below.
Embellishment on the rudimentary life cycle described above evolved in tandem with increasing complexity accompanying multicellularity. For example, in the case of plants, distinct phases include the zygote; post-zygotic development; seed dormancy; germination (often equated with birth); juvenile (vegetative) development; staging of reproduction and longevity as an annual, biennial, or perennial; senescence or lack thereof; and death (Harper and White 1974). Harper (1977, p. 519; see also Niklas and Kutschera 2010) further identifies 10 life cycles among flowering plants with respect to their fertility schedules.
Animals display similarly diverse cycles, among the most dramatic being those of insects with complete metamorphosis. Buss (1987, pp. 165-167) developed seven sets of traits (based on such factors as the nature of the cell enclosure, ploidy, presence of germline sequestration, etc.). If all permutations were possible some 972 life cycles would result; in practice there are specific conditions that influence others and whether or not there can be compensations.
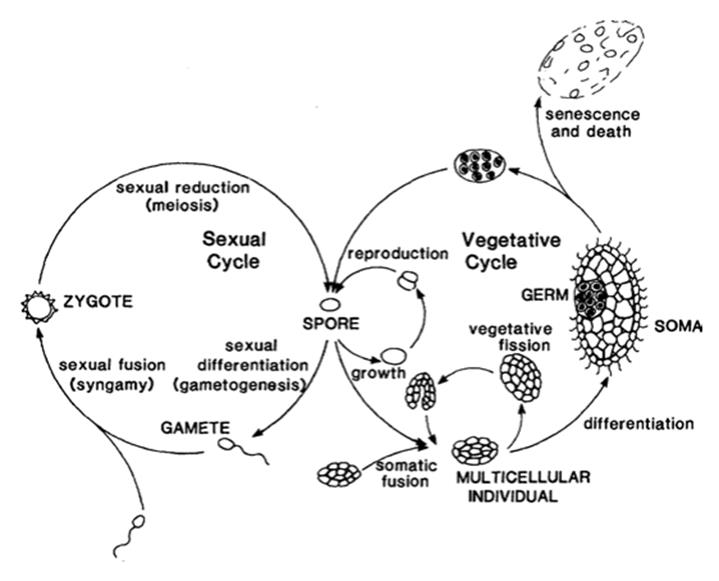
Fig. 6.2. A simplified eukaryotic life cycle, typified here by a protist, showing a sexual phase beginning with a single-celled, diploid zygote and a vegetative phase beginning with a single-celled, haploid spore that links the two phases. Note the seminal role of the unicellular state. From Bell and Koufopanou (1991); reproduced from the Philosophical Transactions of the Royal Society of London B by permission of The Royal Society, ©1991
This reduces the feasible number to a much smaller, though still sizable, subset of a couple of dozen or so and these are based only on the specific conditions chosen by Buss. Clearly, the actual number of life cycles, though numerous, is a subset of the very many theoretically possible, suggesting constraints. A generalized life cycle for multicellular organisms is shown in Fig. 6.2 though there are many variations on the basic theme (e.g., Bell 1994).
Date added: 2025-06-15; views: 342;