A Case Study: On Being a Neighbor
One consequence of a sessile, modular lifestyle is interactions among neighbors. The organism cannot escape from predators by running away or from competitors by moving to a different pasture to graze. Thus, one would expect to find evidence of various defensive mechanisms (and resource exploitation strategies) in such life forms. For instance, competition may be mediated by chemicals, a phenomenon commonly referred to as allelopathy by botanists, and as antibiosis by microbiologists. Sage plants of the genus Salvia in California evidently inhibit the growth of other vegetation at least in part by an allelopathic mechanism (e.g., Muller 1966; for the current status of allelopathy see Inderjit et al. 2011), although interactions are complex and alternative explanations have been put forward (Bartholomew 1970; Orrock et al. 2010).
With respect to microorganisms, the production of antibiotics was formerly alleged to be an artifact of laboratory culture conditions. It is now clear that antibiotics are produced in nature and strong, multiple lines of evidence implicate their role in the antagonistic interaction among some microbes (Raaijmakers and Mazzola 2012). They also have multiple other and perhaps more prevalent roles, among them in intercellular signaling, biofilm ontogeny, virulence, and acquisition of trace elements.
Competitive (and other) interactions are also reflected by morphological behavior. In experiments on competition between the free-floating aquatic plants Lemna and Salvinia, the latter dominated because it overtopped Lemna by expanding its fronds above, and then down onto, the water surface (Clatworthy and Harper 1962). The outcome could not be predicted either by the carrying capacities or by the intrinsic growth rates of the respective species in isolation. When lichens of various morphologies meet on a rock surface, different outcomes are possible, but most commonly the foliose (upright) types override the crustose (appressed) forms (Fig. 5.8; Pentecost 1980). However, the foliose forms are more vulnerable to being removed by abrasion.
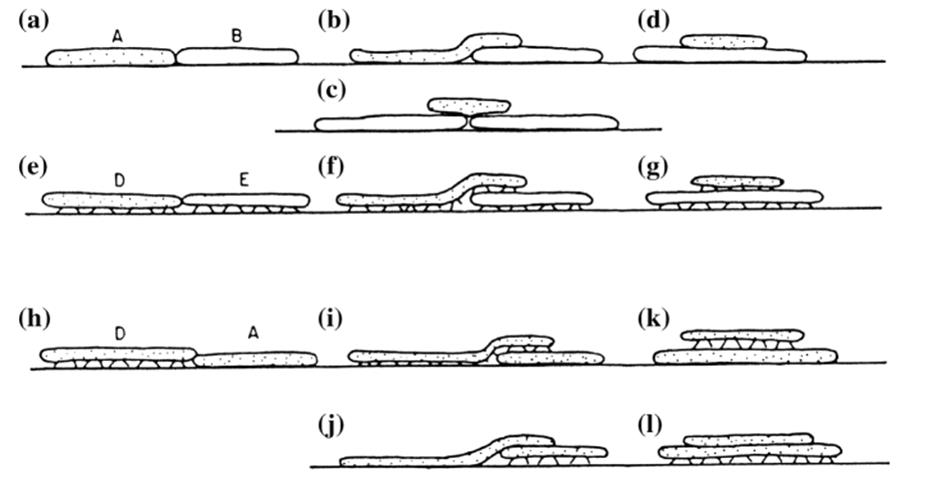
Fig. 5.8. Schematic vertical sections through crustose (mat-forming; shown lying flat like pancakes) and foliose (on stilts) lichens, illustrating the potential range of interactions among these organisms. a-d Interactions between two crustose species, A and B. e-g Interactions between two foliose species, D and E. h-l Interactions between a foliose (D) and a crustose (A) species. Foliose forms can overtop most crustose forms and thus a stalemate (h) or overtopping by a crustose species (j) are uncommon. (d, g, k, l) Designate epiphytic growth which requires colonization of the underlying thallus by propagules containing both the algal and fungal components of epiphytic species, a rather uncommon event. From Pentecost (1980). Reproduced from The Lichenologist by permission of Cambridge University Press © 1980
In the hydroid Hydractinia, intraspecific competition between genetically unrelated colonies also takes on various phenotypic forms ranging from passive to aggressive (Fig. 5.9; Rosengarten and Nicotra 2011). The former response involves the secretion of a fibrous matrix by both colonies and inhibition of growth at the interface. Active rejection is characterized by production of "a specialized organ of aggression," the hyperplastic stolon, at the tip of which nematocysts accumulate and discharge into the neighboring colony. So, as Buss and Grosberg (1990) comment, these simple invertebrates can display a range of aggressive reactions analogous to those of organisms that are much more developmentally and behaviorally complex.
In the fungi, intraspecific contact may be marked by tolerance, fusion (vegetative or somatic compatibility), and potentially the formation of a cooperative ecological unit (physiological individual). This phenomenon has become the standard test for self versus non-self in studies on the distribution of fungal genets. If the interacting colonies differ genetically at more than a few loci, however, the interaction is manifested by a localized darkened zone in which the hyphae are deformed (vegetative or somatic incompatibility). Interspecific responses may occur at a distance or following contact. In the latter case, several outcomes are possible (see Chap. 2 in Cooke and Rayner 1984). Interestingly, overall, the fungal interactions broadly parallel the interactions among hydroids.
An important practical significance of combative responses is that fungi and bacteria can be selected for their competitive, antagonistic ability and used as biological control agents to preempt or counteract plant pathogens. For further information, see Cook and Baker (1983). To cite but one example, the crown gall disease caused by Agrobacterium tumefaciens (overview in Chap. 3) is controlled by a closely related organism, A. radiobacter strain K84, which carries a plasmid coding for the production of an antibiotic that is toxic specifically to virulent A. tumefaciens (Burr and Otten 1999; Brencic and Winans 2005; Gelvin 2010). Similar principles are becoming actively researched and applied to manage the microbial ecology of the human gut (Costello et al. 2012; Donaldson et al. 2016).
Date added: 2025-06-15; views: 365;