A Case Study: When it Helps to Be Small
The plant pathogenic fungi (primarily species of Fusarium, Verticillium, Ceratocystis, and Ophiostoma) that cause vascular wilting are unique in that they set up housekeeping within the water-conducting (xylem) tissues of their hosts. They remain there (and specifically for the most part within the water-conducting elements or xylem vessels) until the late stages of disease development whereupon they may grow into other tissues. Although details of life cycles and pathogenesis vary with each system, there is a common pattern. All these fungi must be able to grow at low oxygen levels within a relatively narrow, segmented pipeline with rough walls, interspersed with obstacles, through which is continuously flowing a dilute nutrient solution.
There are many such pipelines that conduct water from the roots to the uppermost leaves. The 'goal' of the pathogen is to seize the habitat and reproduce. In such an environment the time and resources it would take to produce large spores packed with nutrients would not be advantageous. Rather, selection pressure during the lifecycle phase when the pathogen is residing within the plant's conducting tissues is on numbers of propagules and speed of transport: There is literally a life-and-death race between the host to delimit the infection, and the invader to out-distance the host's response and seize territory. How can the pathogen win the race?
It seems more than coincidental that the wilt-inducing fungi are dimorphic (Puhalla and Bell 1981; see also Nemecek et al. 2006), that is, capable of growing in two distinct forms closely aligned with the parasitic and saprophytic phases of the life cycle. The parasitic phase is typified by single-cell, yeast-like propagules (as bud cells or as microspores borne from simplified stalks or conidiophores) and hyphae, whereas in the saprophytic phase these fungi may produce extensive hyphae and conventional spores, some of which are modified for dormancy or dispersal.
The obvious adaptive value of the yeast or micro-conidial phase in xylem vessels is that fungal biomass is channeled directly into many small cells that move passively upward in the xylem fluid to systemically colonize the host. Numbers may reach about 7000 conidia per mL of xylem fluid (for Verticillium albo-atrum in the hop plant; Sewell and Wilson 1964).
A given length of pipe (technically, vessel) rarely extends the full length of the plant and water must pass through a thin cell wall (pit membrane) as it moves laterally from one vessel to the overlapping portion of the next. These walls block passage of spores. Vessels, in turn, are composed of many segments of pipe (vessel elements) stacked one upon the other. The end walls (perforation plates) where the segments join are sufficiently open that water flow is not impeded appreciably but, depending on species of plant, rather than containing one large opening may consist of many smaller ones that entrain fungal propagules.
The common strategy of the wilt fungi is to release into the sap stream abundant spores that travel for a distance before becoming obstructed; they then germinate quickly to produce hyphae that grow through the obstruction, produce another round of yeast-like cells or microconidia, and the sequence is repeated. Thus ensues a pitched battle between the host—which attempts to restrict the invasion by various mechanical and biochemical mechanisms—and the pathogen, which must advance rapidly if it is to escape the counterattack. It does so in successive waves of propagule release followed by growth through obstacles, much like an infantry battalion advances to secure successive beachheads. This battle is illustrated in Fig. 4.13.
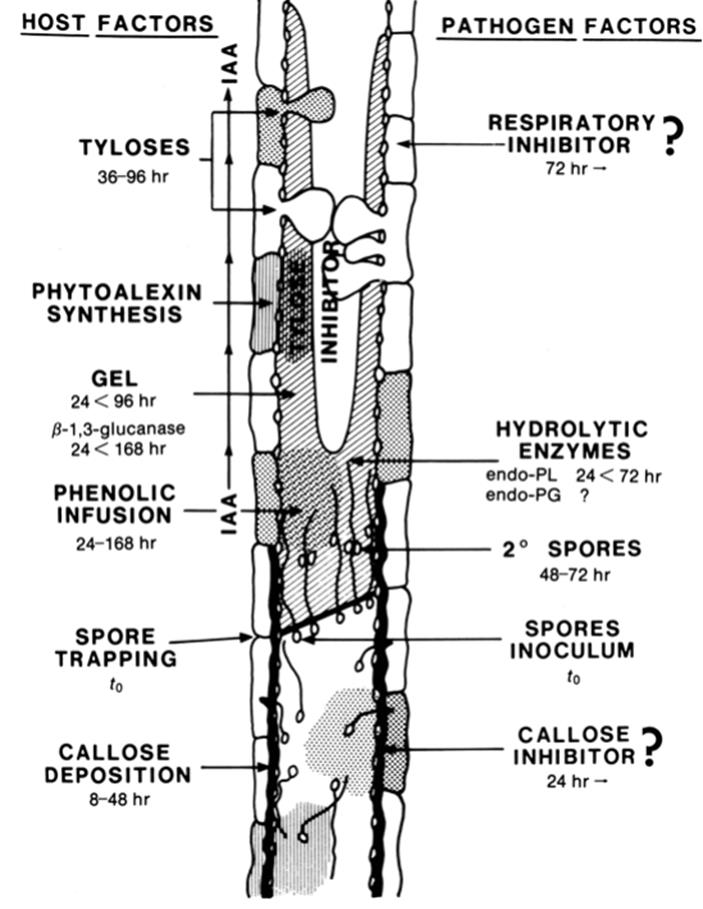
Fig. 4.13. Model showing in longitudinal view how the plant host and vascular wilt pathogen confront each other in a portion of the water-conducting (xylem) tissue. Events shown occur over the initial 7-day time period after infection; a race ensues between the pathogen, which produces factors to promote systemic colonization; and the host, which attempts to physically and chemically contain the pathogen. The large central 'pipeline' is a portion of a vessel showing many small pathogen spores, some of which are entrained at a vessel element end wall (perforation plate), subsequent invasion of the adjacent vessel element by spore germination, followed by secondary rounds of sporulation. Spores are small and easily moved upwards in the transpiration stream though impeded by obstacles produced by the host. The difference between a resistant and susceptible plant depends critically on speed or response of the host. Tyloses = balloon-like invaginations of parenchyma cells adjoining the vessel elements into the lumen of the vessels; IAA = plant hormone indoleacetic acid; phytoalexin = specific type of inhibitory chemicals elucidated by host to inhibit pathogen. Reproduced from Beckman, C.H. 1987. The Nature of Wilt Diseases of Plants, by permission of the American Phytopathological Society, St. Paul, MN, ©1987
A related and interesting side note: wilt diseases are classically a problem of Angiosperms and not Gymnosperms because of the anatomical difference in their water-conducting elements (Dimond 1970). For additional information, see Esau (1965), Mace et al. (1981), Pegg (1985), and Evert (2006).
Date added: 2025-06-15; views: 304;