Characteristics of Amphibians and Reptiles. The third part
From the foregoing, it is evident that both birds and mammals have evolved from the group that we refer to as reptiles. Thus, the Class Reptilia, excluding birds and mammals, is not a real evolutionary unit since it incorporates elements that are more closely related to groups that are not regarded as reptiles than they are to other nominal reptiles (Fig. 2.1). Even at the level of living "rep- tiles," this problem applies. For example, the crocodylians are more closely allied to birds than they are to lizards and snakes, yet birds are not usually regarded as reptiles.
The currently accepted pattern of relationships among all of these forms is represented as a cladogram (a type of genealogical tree) in Fig. 2.1. In this book, we will also exclude birds from our consideration. This poses some prob- lems from the point of view of generalizing about living reptiles or comparing them to one another. These problems do not apply to the living lissamphibians as evidence points to their common ancestry, independent of other fossil and living groups.
The living reptiles have in common: a scaly, keratinized integument; true claws; a cleidoic egg; extraembryonic membranes; modified wrist and ankle joints; full complement of cranial nerves; internal fertilization following copulation; and excretion of uric acid as the primary nitrogenous waste product. One of the most crucial developments in reptilian phylogeny was the evolution of the cleidoic egg (Fig. 2.6).
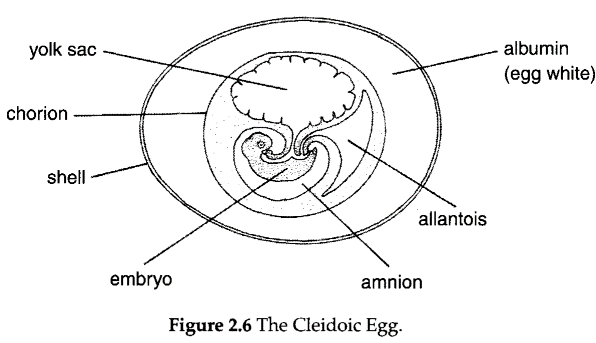
Cleidoic eggs, for example chicken eggs, have controlled exchanges with the environment, allowing for the diffu- sion of respiratory gases while protecting the developing embryo from desiccation and mechanical shock. Such eggs are able to develop terrestrially, but can only do so if deposited in situations with particular environmental characteristics. Typically, reptiles do not incubate their eggs, and thus selection of the nest site is critical. This is even more apparent for those reptiles that have parchment-shelled (pliable and porous) rather than calcareous-shelled (hard, as in chickens) eggs. There are three extraembryonic membranes typical of reptiles, birds and mammals (the amniotes), and these play critical roles in development: the amnion provides a fluid-filled sac around the embryo; the allantois serves as a site of storage of waste products and, along with the chorion, provides a surface for respiratory gas exchange.
The second reptilian "innovation" was a less permeable integument. This, however, reduced the potential for gas exchange through the skin, but permitted the reptiles to fully exploit the terrestrial realm. Crucial in this transition was an effective waterproofing of the outer layers of the skin that greatly reduced the osmotic flux potential, more effectively isolating the internal environment of the body from the external environment. This same modification was inherited from the reptiles by both birds and mammals.
The reptiles, like the amphibians, are ectothermic (obtaining their body heat from external sources) and poikilothermic (having a fluctuant body temperature). They are thus unable to fully regulate their body temperature physiologically. They do, however, thermoregulate by behavioural means (Fig. 2.7).
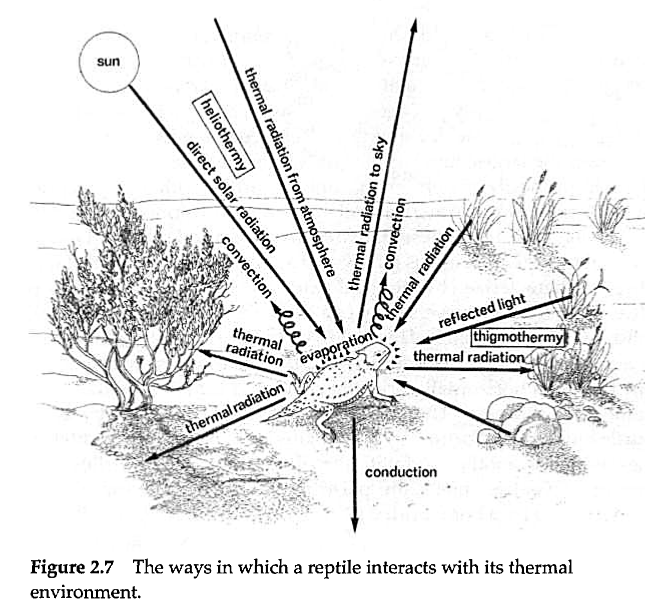
Those which gain heat directly from the sun's rays are known as heliotherms, and those that gain it from the ground upon which they rest are known as thigmotherms. While reptiles in general may require a fairly high environmental temperature to maintain them in an active state, they are very susceptible to overheating. They may be killed when the body temperature rises to a level only a few degrees above that to which they will normally expose themselves. Body temperatures of about 46°C may be potentially lethal to certain desert lizards, whilst temperatures of about 40° may be tolerated without stress.
The germ cells (those associated with reproduction), however, may have a higher sensitivity than the body in general, and males of some lizards become sterile at temperatures which apparently cause no physical discomfort. Susceptibility to heat is probably in large part due to the fact that reptiles have no rapid cooling device comparable to the sweat glands of mammals. Many species do pant, but this is a drastic solution and rapidly depletes body water. Postural thermoregulation is also possible, and many forms stand high on their legs in order to radiate heat. Behavioural thermoregulation ensures that body temperatures remain within a certain tolerable range, known as the preferred tem- perature. This is basically higher in diurnal (day active) than it is in nocturnal (night active) and burrowing forms.
Some physiological modifications to control temperature also exist. In large active lizards, such as monitors, the rate of heat loss or absorption is influenced by changes in distribution of circulating blood. This distribution is between the superficial and deep tis- sues of the body. Colour change is another method of temperature control, and the amount of heat absorbed by an object depends to some extent upon its coloration. Thus, some species tend to grow darker when they are first exposed to the sun, in order to increase heat absorption, and then become lighter, in order to retard further temperature rise.
Reptiles are more resistant to extremes of cold than they are to extremes of heat. Prolonged exposure to sub-zero temperatures is probably lethal, but above this level many species can exist for long periods in a state of torpor. In temperate zones, such torpor is always a feature of reptile life, as it is of amphibian life. These organisms thus become adapted to a seasonal climate in which certain parts of the year provide too little warmth for them to remain active. Many forms of both amphibians and reptiles in cold regions tend to burrow beneath the permanent frost line in the winter to avoid extreme temperatures.
As a rule, reptiles breathe atmospheric air and utilize lungs. Gills are never present. The pulse pump seen in the frog, in which air is forced into the lungs from the buccal cavity, is replaced in reptiles by the suction pump. This involves an interaction between the ribs, the viscera and many muscles to enable the thorax to expand and contract. This suction pump has advantages over the force pump. The lungs are filled or emptied in a single pass, independent of the size of the buccal cavity. In frogs, only as much air as can be housed in the buccal cavity can be taken into the lungs at any one time. The flow of air in reptiles thus becomes more or less continuous and the buccal cavity, freed from its role in ventilation, can be designed more effectively for food capture and processing. It also enables the skin to become armoured as a protective device and keratinized to prevent water loss, as the skin is no longer used as an organ of gas exchange.
The 6,970 species of living reptiles are divided into five major groups, each characterized by a different body form and different biological characteristics. The relationships of these groups to each other are indicated in Fig. 2.1.
There are more than 250 species of living turtles (Fig. 2.8).
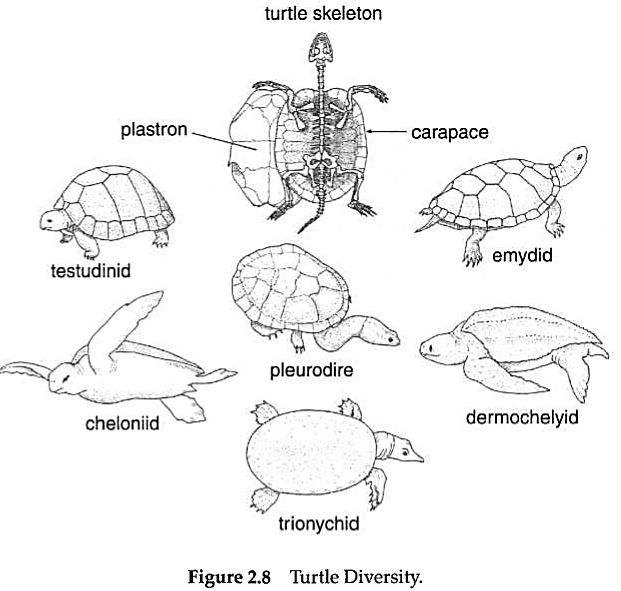
This number includes both fresh-water and marine turtles, as well as partially and wholly terrestrial forms. Turtles, the Testudinata (or Chelonia), are of course immediately identified by their "shells," consisting of a dorsal carapace and a ventral plastron. In addition, turtles exhibit the loss of teeth; the absence of temporal fenestrae; the placement of the shoulder girdle inside the rib cage; the modification of the neck vertebrae for packaging inside the shell as the head is withdrawn; the presence of a single copulatory organ in males; and modification of ventilatory motions in association with the presence of a rigid shell. Turtles range in size from less than ten centimetres in carapace length for some testudinids to more than 2.0 metres in length and 800 kg in weight for Dermochelys coracea, the leatherback sea turtle. As a group, turtles occur worldwide, except for high latitudes and some remote oceanic islands.
Date added: 2022-12-11; views: 913;