Fishes and Amphibians in Temporary Waters: Adaptations and Ecology
Fishes. Apart from highly specialized forms, such as lungfishes which can aestivate in the bottom mud during the dry phase, fishes tend to be absent from temporary ponds. Temporary streams, on the other hand, may support fish populations of considerable size and diversity. This is the result of either some streams becoming intermittently connected to permanent waters from which fishes can migrate, or fishes surviving in permanent pools left in some drying streambeds. Studies of the ecology of fishes in temporary streams are, however, few in number. Possibly, this is because although large rivers do occasionally dry up, particularly in the tropics, intermittency, particularly in temperate regions, is more a characteristic of smaller bodies of water that usually support populations of fishes of little economic or recreational importance. There does exist, however, a more substantial literature on those fish species that use temporary pools on the floodplains of large rivers during part of their life cycles; these will be discussed in more detail in Chapter 8.
Three species of fish are known from the Kalahari Desert. Clarias gariepinus is a catfish that possesses suprabranchial respiratory organs. The other two species are both cichilds, Tilapia sparrmanni and Hemihaplochromis philander; all three species can survive in very little water as stunted individuals (Cole 1968).
In North America, out of a total of 50 species of fish found by Williams and Coad (1979) in the Grand River watershed, Ontario, only 12 were collected from three intermittent tributaries. Species in the latter were largely members of the families Cyprinidae (minnows) and Percidae (perches), with one species from each of the Catostomidae (suckers) and Gasterosteidae (sticklebacks). The intermittent streams entirely lacked catfishes, sunfishes, and salmonids. The brook stickleback Culaea inconstans and the cyprinids Pimphales notatus and P. promelas showed physiological tolerance in that they survived poor water quality, high temperatures, and crowding in shrinking summer pools. Catostomus commersoni (white sucker) and Semotilus atromaculatus (creek chub) on the other hand, moved into the tributaries, spawned, and then left. The main advantages to fish species colonizing intermittent streams appear to be plentiful food, earlier spring breeding (as the water is often warmer than in adjacent permanent streams), and reduced predation by large fishes.
A problem faced by fishes moving into intermittent streams is that they may become stranded and die if the pools dry up completely. The longfin dace, Agosia chrysogaster, possesses behavioural adaptations that contribute to its success as the only species to consistently use intermittent streams in the Sonoran Desert of Arizona. These streams dry up to form pools separated by lengths of dry streambed. The fishes position themselves in the current and this minimizes the chances of them becoming stranded by falling water levels. This species quickly invades new habitats during wet periods when flow is continuous and is capable of existing for at least 14 days in areas where there is no free water, provided that there is moisture beneath mats of algae (Minckley and Barber 1971; Bushdosh 1981). Avoidance of the stream edges and shallows reduces predation from birds and mammals. In contrast, high predation pressure from large aquatic and terrestrial predators has been put forward to explain the coexistence of three catfish species in swamps in Suriname. Mol (1996) suggested that as drying swamp pools become more restricted, predators, such as caiman and birds, exert so much pressure on populations of Hoplosternum littorale, H. thoracatum, and Callichthys callichthys that none can dominate.
Another fish species that is adept at surviving in intermittent streams is Poecilia reticulata, the guppy. Although more frequently studied in the clear water, upland permanent streams of the Northern Range of its native Trinidad, populations thrive in the lowland rivers of the southwestern part of the island. During the dry season, hundreds of individuals survive in shallow (5-10 cm), highly turbid, streambed pools no more than 1 m or so in diameter, where they appear to survive by respiring the thin surface layer of oxygen-rich water (Alkins-Koo 1989/90). Alkins-Koo's study focused on two intermittent streams on the Chatham Peninsula: the Carlisle and Quarahoon rivers, both of which range from 1 to 8 m wide and up to 3 m deep in the rainy season, to isolated or chains of pools in the dry season. From 1980 to 82, she collected 31 species from these two rivers (Table 4.5), which, despite the relatively small drainage area and the interrupted flow regime, represented almost half of the freshwater fish fauna of the island. Alkins-Koo suggested that this high diversity may have resulted from several habitat features: (1) an intermediate level of disturbance, resulting from the annual flood-drought cycle (see Townsend et al. 1997); (2) the presence of extensive pool refugia; and (3) the dynamic state of the local fauna on the peninsula, due to continued colonization from the nearby mainland of Venezuela. In terms of adaptations to surviving in these two streams, Alkins-Koo pointed out that many of the species are known to be able to endure fluctuating environmental conditions, especially stagnation and hypoxia, increased predation, and crowding. Several species are capable of breathing air, and Rivulus hartii, C. callichthys, and Synbranchus marmoratum, for example, can move overland, allowing them to colonize new habitats. In particular, S. marmoratum can survive without free water, in burrow systems (Kramer et al. 1978).

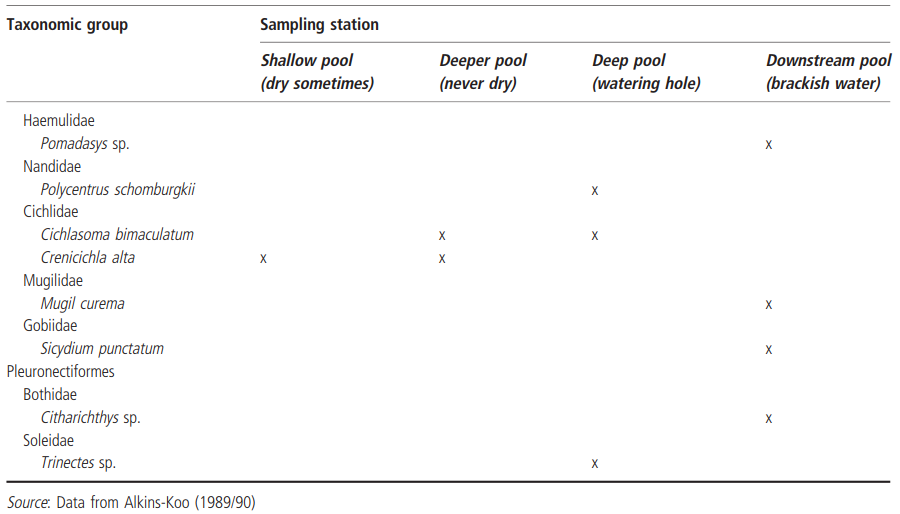
Table 4.5. Teleost fish species collected from two intermittent streams on Trinidad, West Indies
Survival and between-pool migration of Clarias anguillaris on the floodplain of the Sokoto-Rima River, in Nigeria, was also possible by this species' ability to breathe air. However, Hyslop (1987) proposed that its highly varied diet (from higher plants and algae to invertebrates and other fishes) also contributed to its success in temporary waters. Ouboter (1993) summarized some of the other adaptations that allow fishes to survive in freshwater swamps in Suriname; these are summarized in Table 4.6.
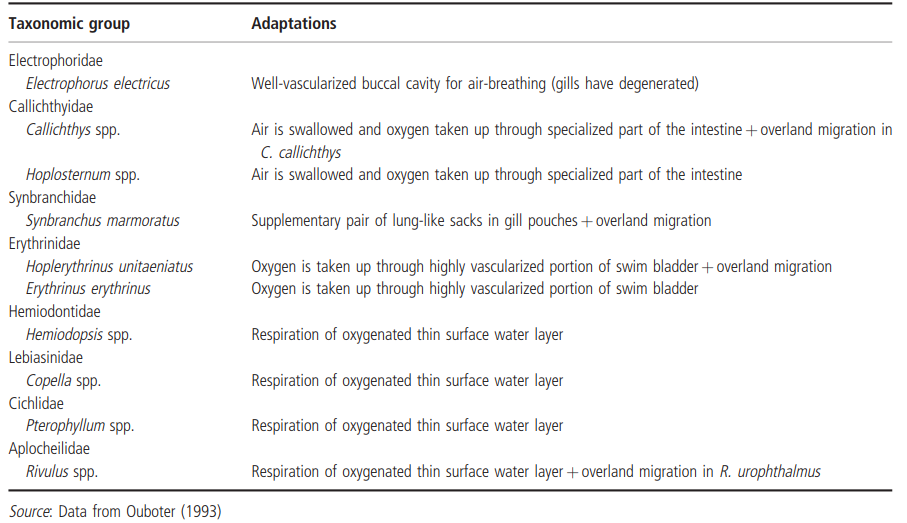
Table 4.6. Summary of some of the adaptations that allow fishes to survive in the freshwater swamps of Suriname
The capacity of temporary water fishes to air- breath is currently causing some concern in the United States. An established population of airbreathing snakeheads (Channidae: Channa marulius) was discovered in Broward County, Florida, in 2000, and introduced populations have been captured in several other states (Courtenay and Williams 2005). Arising from Asia, these fish are voracious feeders with a diet that, in the United States, now includes native fish species. Snakeheads possess suprabranchial chambers for breathing air, combined with a ventral aorta that is divided into two parts that permit bimodal (air and water) respiration. Airbreathing allows chan- nids to be very adept at dispersing among ponds via overland travel, creating the potential for displacement of native fish species.
Amphibians. Frogs and salamanders are common inhabitants of temporary ponds and cosmopolitan genera include Rana, Hyla, and Ambystoma. Species richness has been found to be highest in habitats that exhibit an intermediate level of spatial-temporal variability. In the Rhone Valley, for example, these habitats are mesotrophic temporary ponds, and species align themselves along a variability gradient (Joly and Morand 1997). The most unstable sites are colonized by the yellow-bellied toad, Bombina variegata. Bufo calamita, Hyla arborea, and Pelodytes punctatus (the parsley frog) are characteristic of ponds of intermediate stability. Where water levels are more predictable, Bufo bufo, Rana temporaria, and R. lessonae occur. Salamandra salamandra and Alytes obstetricans (the midwife toad) are absent from the floodplain, probably because the adults cannot tolerate submersion during floods. Species such as R. dalmatina, R. ridibunda, and Triturus helveticus are ubiquitous, able to breed in ponds spanning the gradient.
Clearly, the aquatic larvae of all these species are severely threatened by any untimely onset of the dry phase of the habitat. Wilbur and Collins (1973) have suggested that an endocrine- controlled, metabolic-feedback mechanism exists in temporary pond species. Should the rate of larval growth be slow, metamorphosis to the adult stage is initiated once a certain minimal larval size is attained. Although the resulting small adult may face disadvantages in the terrestrial environment, these are less than those facing the larva if the pond dries up prematurely. If, however, the larval body size is small but its rate of growth is fast, metamorphosis is delayed so as to maximize the animal's growth potential in the pond. Control of metamorphosis is thus related to the stability of the habitat and species with a fixed size for metamorphosis are therefore excluded from temporary waters. In Western Australia, all the species of Heleioporus (burrowing frogs) breed in the winter and lay their eggs in a frothy mass in a burrow dug by the male. The site chosen is always one which will be later flooded by heavy winter rains.
Species of Pseudophryne (toadlet frogs) have similar egg-laying habits. As the rain raises the level of the water table in the burrow, the larvae break free of the egg mass and develop to metamorphosis often in no more than 0.5 l of water. In the two western species of Pseudophryne, larval development takes slightly longer than 40 days. The aquatic phase of their ponds seldom lasts more than 50 days so, potentially, there is little leeway. This is offset, however, by the ability of the eggs to complete embryonic development (6-8 days) in the absence of free water, so that the larvae are ready to hatch as soon as the rains come. In the event of a delay in rainfall, hatching can be postponed for several weeks (Harrison 1922; Main et al. 1959).
Adults of the five species of Australian Heleio- porus, together with those of the genus Neoba- trachus (spadefoot and other frogs), can withstand a drop in the water content of their bodies of up to 45% of their body weight. Rehydration rates in species of Neobatrachus vary according to the severity of water loss in their particular habitats. For example, species that live in the arid interior rehydrate faster than those from the wet coastal regions of the southwest. In contrast, Heleioporus shows no difference in rehydration rates among species spanning a wide spectrum of aridity. It is thought that because all species of Heleioporus are superior burrowers, selective pressures for increasing the speed of rehydration may not operate (Bentley 1966).
Burrowing seems to be a common method of surviving droughts in amphibians, as even the ability to get just a few centimetres below ground level places the animal away from the drying effects of sun and wind, and into a more moist environment. Even in deserts, moisture from past rains can remain trapped for years in sand at depths of only 20-30 cm (Bagnold 1954). Scaphiopus couchi, the spade-foot toad of California aestivates in its burrow surrounded by a layer of dried, skinlike material. This may help to limit evaporation of moisture from its body in much the same way as the cocoon-like covering of the African lungfish. Another species, S. hammondi, lines its burrow with a gelatinous substance that presumably slows down water loss. In this genus, aggregations of tadpoles have been found shortly before metamorphosis and subsequent emergence from temporary ponds (Bragg 1944). It has been suggested that these dense aggregations may conserve water, as the combined rapid beating of many tadpole tails tends to deepen that part of the pond basin and water from shallower parts of the pond will drain into it.
That amphibians, particularly frogs and toads, can successfully contend with the intermittent availability of water (though many people think of them as being associated with cool, moist environments) is evidenced by phenomena, such as those that occur in the western deserts of Australia. Here, after rain, the number of frogs emerging from burrows is so large as to interfere with rail transportation, as thousands of frogs are crushed as they attempt to cross railway lines thus making traction impossible (Bentley 1966). Colonization of new ponds by the alpine newt, Triturus alpestris, has been shown to be by both adults and juveniles (Joly and Grolet 1997).
Mayhew (1968) summarized the general adaptations of amphibians found in dry areas as follows:
- no definite breeding season;
- use of temporary waterbodies for reproduction;
- breeding behaviour initialized by rainfall;
- loud voice in male attracts both females and other males, resulting in rapid congregation of breeding animals;
- rapid development of eggs and larvae;
- omnivorous feeding habits of tadpoles;
- production of inhibiting substances by tadpoles which influence the growth of other tadpoles;
- high tolerance of heat by tadpoles;
- adults have metatarsal 'spades'for digging;
- ability to withstand considerable dehydration, compared with other anurans;
- nocturnal activity.
A particularly noteworthy observation on amphibian populations in temporary waters is that there exists a great deal of variation in reproductive characteristics (especially in egg and clutch size) among individuals. Kaplan (1981) has examined this variation in the light of theory on 'adaptive coin-flipping' or natural selection for random individual variation. Given two genotypes with equal mean fitnesses, the genotype with less variance in fitness will eventually outcompete the genotype with more variable fitness (Felsenstein 1976). Kaplan argues that natural selection for random individual variation has been overlooked because, in general, neo-Darwinian theory is a theory of genes and not a theory of development and, consequently, population geneticists often do not take into consideration influences of the environment on development. A well-buffered phenotype may be advantageous in many cases, but a less-well-buffered developmental system also might be of advantage to an individual, particularly in environments that are temporally variable. This is supported by the observations made on variation in size for the onset of metamorphosis, discussed earlier. Further evidence for the selection of phenotypic plasticity in amphibian populations living in highly variable environments has been provided by Van Buskirk (2002), although he concluded that species-specific attributes may sometimes obscure this relationship. Van Buskirk also suggested that behavioural solutions to surviving in such environments are likely to evolve under different scales of variation than morphological responses.
In a large-scale experiment, Wilbur (1987) demonstrated that when several anuran species coexist, assemblage structure is determined by an interaction between biological and environmental factors. In an assemblage comprising Rana utricu laria, Scaphiopus holbrooki, Bufo americanus, and Hyla chrysoscelis, neither predation nor competition proved to be the single unifying force, instead they interacted in determining the consequences of the date of pond drying to the emergence success of each species. In another experiment, Blaustein et al. (1996) found that Hyla savignyi and Bufo viridis were heavily preyed upon in ponds containing larvae of the fire salamander S. salamandra infraimaculata. Interestingly, the tadpoles tended to be larger in the pools with salamanders. Not only did this predator affect the anuran populations, but it had an impact on most of the other pond inhabitants. For example, it reduced invertebrate species richness by 53%, eliminated the large cladoceran, Simocephalus expi- nosus, and reduced populations of the numerically dominant calanoid copepod, Arctodiaptomus similis, and a species of Chironomus. As a consequence of reduced invertebrate grazing pressure together with increased nutrient input from the salamander's excreta, periphyton and bacterial populations rose. Laurila and Aho (1997) examined whether adult behaviour of R. temporaria might play a role in larval survival through selection of ponds that are predator free. However, there was no evidence to support this, leading these authors to conclude that competition, in combination with pool desiccation, was the main factor affecting fitness, perhaps favouring females that primarily selected vacant pools, or pools with low densities of competitors.
Temporary Wetlands as Vital Habitats for Reptiles, Birds, and Mammals
Reptiles. A variety of terrestrial reptiles use temporary waters as sources of drinking water. In addition to these, there are several types of reptile that live in close association with both permanent and temporary waterbodies, for example, crocodiles, monitor lizards, turtles, and iguanas. Alkins-Koo (1989/90) recorded four species of reptile from two intermittent streams in southwestern Trinidad: the turtles Phrynops gibbus, Rhinoclemmys p. punctularia, and Kinosternon s. scorpioides, and the caiman Caiman crocodilus. In the Amazon basin, the breeding cycle of the latter appears to be closely linked to fluctuations in water level, with nests constructed in the grass mats that cover the margins of large lakes (Lang 1989). Three species of caiman have been recorded in the Pantanal wetlands of Brazil: Paleosuchus palpebrosus and Caiman latirostris (though both may be now locally extinct), and C. yacare, the jacare, which is still abundant. The jacare frequents shallow bays, where it hauls out during the day. Its food consists primarily of aquatic snails and small fishes, but aquatic insects, especially beetles and hemipterans, are important in the diet of juveniles (Por 1995). The Nile crocodile, Crocodylus niloticus, is known to use temporary streams that flow in the rainy season as dispersal routes to new permanent water bodies.
Ehrenfeld (1970) found that alligator holes (temporary ponds excavated by the reptiles) in the Florida Everglades serve as collecting ponds and biological reservoirs for the surrounding aquatic life—both vertebrate and invertebrate—in the dry season. Rich growths of algae and higher aquatic plants are nourished by the reptiles'droppings and these, in turn, maintain a variety of animal life. At the end of the drought, the survivors move out to colonize the glades anew. The Florida Everglades also support a large number of snakes and several lizards that commonly use temporary pools and marshes as habitats in which to feed, drink, or regulate their body temperature; these include the following: Eumeces inexpectatus (southeastern five-lined skink), Ophisaurus ventralis (eastern glass lizard), O. compressus (island glass lizard), Nerodia floridana (Florida green water snake), N. taxispilota (brown water snake), N. fasciata (Florida water snake), N. clarkii (mangrove salt marsh snake), Seminatrix pygaea (South Florida swamp snake), Thamnophis sauritus (peninsula ribbon snake), Regina alleni (striped crayfish snake), Farancia abacura (eastern mud snake), Opheodrys aestivus (rough green snake), Drymarchon corais (eastern indigo), Elaphe obsoleta (Everglades rat snake), and Aghistrodon piscivorus (Florida cotton- mouth) (Steiner and Loftus 1997). Such a high diversity of predators must have a significant effect on the wetland communities. In the water hyacinth- choked savanna wetlands of Venezuela, anacondas (Eunectes murinus) prey on capybaras and wading birds that frequent these waters. These reptiles also are highly dependent on these habitats for other life processes, including mating (breeding balls), dispersal, and thermoregulation (Figure 4.17).

Figure 4.17. A female anaconda (~4.5 m long) living in the water hyacinth-choked savanna wetlands of Venezuela (photograph provided and copyrighted by A. Chartier)
Birds. Wetlands are crucial habitats, in terms of both reproduction and feeding, for many bird species throughout the world. Indeed, a number of studies have demonstrated a close correlation between the annual production of waterfowl and the number of wetlands. For example, breeding densities and brood numbers of the mallard (Anas platyrhynchos) were found to be highest in years when the densities of ponds holding water were highest (Leitch and Kaminski 1985). Poiani and Johnson (1991) have pointed out that the quality of habitat for waterfowl is highly dependent on the mix of permanence types found in wetland complexes. For example, temporary pools provide an abundant food source early in the season, and tend to be heavily used by dabbling ducks in the spring. Seasonally flooded wetlands also provide nesting habitat and sites for brood-rearing for both dabbling and diving ducks, especially in years of high water. As wetland areas dry, birds capable of re-nesting tend to relocate to open-water areas (Swanson 1988).
In Australia, extensive breeding of waterfowl occurs in floodplain areas adjacent to rivers. In lightly wooded and treeless plains, vast areas of new waterfowl habitat are created when rivers overflow their banks. These shallow waters are soon colonized by huge numbers of grey teal, pink-eared ducks and shovelers, and by lesser numbers of black duck and white-eyed duck (Frith 1959; Timms 1997). Piscivorous bird species also have life cycles closely linked to floods such that, in Africa, fledglings are produced just when small fishes appear in floodplain pools. There is also heavy predation as the floodwaters recede and fishes become stranded in temporary pools and channels (Bonetto 1975). Foraging success of bald eagles (Haliaeetus leucocephalus) has been shown to be strongly dependent on fluctuating water levels of the Colorado River, Arizona, being higher at low flow, particularly in isolated pools (Brown et al. 1998).
In Asia, wetlands are extremely productive in terms of invertebrates, fishes, and amphibians, and this food base draws in both resident species, such as storks and herons, and migrants, such as waders and egrets. The wetlands of northern Asia, in general, and Siberia, in particular, are among the most extensive in the world, and regions, such as the Ob floodplain, are especially important breeding grounds for water birds. Not all wetlands are natural, however. In the nineteenth century, for example, the Maharaja of Bharatpur created what has become one of the finest water-bird sanctuaries in the world. Originally intended as a shooting preserve, its construction involved building small dykes and dams, and diverting water from an irrigation canal. Within a few years, this 29 km2 low lying area became surrounded by marginal forests, and the complex now supports over 300 species of bird. Among these is the Siberian crane (Grus leucogeranus) which flies over 6,400 km from its summer retreat to get there.
Of course, birds also provide some benefits to wetlands, primarily in the form of nutrient input. For example, Lesser snow geese were measured as increasing the nutrient loading rates in wetland ponds in New Mexico by up to 40% for total nitrogen and 75% for total phosphorus; nitrogen proved to be consistently limiting to algal primary production in these ponds (Kitchell et al. 1999). Many bird species are also the primary dispersal mechanisms for temporary water invertebrates and plants, which sometimes show distribution patterns that coincide with migration routes.
Mammals. Temperate wetlands tend to support relatively few species of mammal. Those present either eat fishes and amphibians, like otters and mink, or feed on wetland vegetation, like mice, rats, beavers, and water-voles. Unfortunately, information on mammals frequenting isolated waterbodies, per se, seems rare. However, in a study of the use of a vernal pool by small mammals in chaparral and coastal sage scrub communities in southern California, Winfield et al. (1981) found that it did not appear to be used heavily. This was despite the fact that it provided a potential source of food in the form of protracted growth of riparian vegetation and semi-aquatic, ground-dwelling insects. Of seven common species of mouse, rat, and rabbit, only one, Reithrodontomys megalotis, the western harvest mouse, had a higher estimated population at the pool than elsewhere, but these results were considered tentative.
In contrast, wetlands in southern Asia have a high diversity which includes some rare species restricted to these habitats, such as Lutra sumatrana (hairy-nosed otter), and the wholly aquatic dugong and Chinese river dolphin. Table 4.7 lists some other mammal species commonly associated with Asian wetlands.
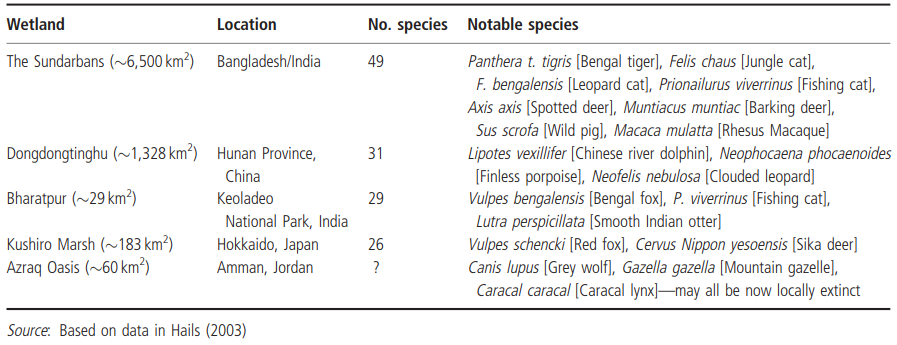
Table 4.7. Mammal species associated with a variety of Asian wetlands
In Africa, many species of wildlife move onto the floodplains of rivers during the dry season in search of grazing and prey. Some antelopes, such as the bushbuck, Tragelaphus scriptus, and the lechwe, Kobus leche, migrate back and forth across swampy ground as the floods rise and fall. Their life cycles are aptly timed such that they drop their young as the floodwaters recede and new pasture is exposed (Welcomme 1979). Other African antelope that live in wetlands include the sitatunga (Tragelaphus spekii) and the water chevrotain (Hyemoschus aquaticus), both of which are often seen almost fully submerged in water. In addition, the waterbuck (Kobus ellipsiprymnus) is a shaggy antelope with oily fur that feeds around wetlands.
Hippopotamus are important transporters of nutrients through their habits of grazing on floodplains at night and depositing large quantities of nutrient-rich dung in water as they bathe during the day. The capybara, Hydrochoerus, of South America similarly inhabits floodplains where it feeds on grasses and aquatic plants; it is generally associated with permanently wet areas (Gonzales-Jimerez 1977).
In Kenya, some water holes are formed from the erosion of termitaria by wildlife such as rhinoceros and hartebeest rubbing against the mounds. When subsequently weathered below soil level, water collects in them and elephants, warthogs, buffalo, and other animals use them as sources of drinking water, thus accelerating the drying-out process (Ayeni 1977). In many arid or semi-arid regions of the world, scattered water holes provide drinking water for animals that can move long distances. Most large mammals in areas such as the grasslands of Africa generally drink every one or two days, in hot weather, and are thus very dependent on finding waterbodies regularly. Only a few species of ungulate are capable of going without water for longer periods (e.g. camels), although there are a number of rodents (e.g. kangaroo rats, gerbils, pocket mice, jerboas) that seem capable of existing on water obtained from food alone.
Date added: 2026-07-14; views: 7;