Crustaceans in Temporary Waters: Ecology and Adaptations
Crustaceans. Crustaceans are an ancient group dating back to marine environments of the lower Cambrian era. Of the approximately 40,000 extant species known, about 10% live in inland aquatic habitats. The latter include some extreme habitats, such as polar ponds and streams, hotsprings, highly saline lakes, as well as a wide range of intermittent and episodic waters. For convenience, coverage here will subdivide crustaceans into three main groups: the large, but primitive Branchiopoda; the large, more advanced Malacostraca; and the microcrustaceans.
In terms of diversity, temporary ponds are frequently highly species-rich, especially in their zooplankton. For example, ponds on the upper coastal plain of South Carolina support 44 species of cladoceran and 7 species of calanoid copepod (Mahoney et al. 1990). In a single temporary pond in western Morocco, Thiery (1991) found six species of Anostraca, two of Notostraca, and two of Conchostraca (Spinicaudata). Petrov and Cvetkovic (1997) similarly found up to seven species of large branchiopod in a single pond in Yugoslavia. Eder et al. (1997) provided evidence for the temporal segregation (related to hydrology, temperature, and water chemistry requirements) of branchiopods in Austrian waters: the anostracans Chirocephalus shadini and Eubranchipus grubii, and Lepidurus apus (Notostraca) occurred in late winter and spring; Branchinecta ferox, B. orientalis (Anostraca), Cyzicus tetracerus (Conchostraca), and Chirocephalus car- nuntanus occurred only in spring; the anostracans Streptocephalus torvicornis and Tanymastix stagnalis, and Eoleptestheria ticinensis (Conchostraca) were present in spring and summer; and Branchipus schaefferi (Anostraca) was found only in summer and autumn.
Despite there being limited comparative data on temporary water crustaceans, properties of the Branchiopoda, and one of its orders in particular— the Anostraca (brine shrimp and fairy shrimp), are better known. Anostracans are important members of the communities of temporary lentic waters throughout the world (Table 4.3).
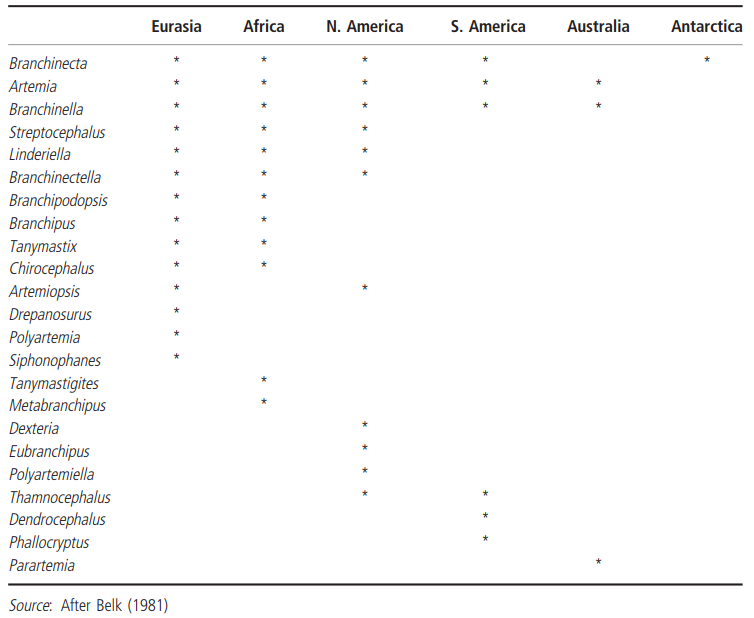
Table 4.3. World distribution of the 23 genera of Anostraca
The majority (17) of present-day genera (23) appear either to have evolved since the formation of the modern continents or represent relict groups. Resolution of these alternatives is hampered by the paucity of their fossil record (Tasch 1969). Modern distribution patterns that suggest extensive distribution in Pangea (the single super-continent that existed in the Paleozoic era, some 200 million years ago) are seen in only three genera—Artemia, Branchinella, and Branchinecta. Three other genera, Artemiopsis, Linderiella, and Streptocephalus, have current distributions that suggest that they were widely distributed across Laurasia (the northern landmass that was derived from the breakup of Pangea in the late Triassic era, some 180 million years ago; Figure 4.8). Belk (1981) proposed that many of the presumptive Pangea genera do not now occur on all of the modern continental fragments of Pangea because of ecological factors. Branchinecta, for example, is a genus of cold-water adapted species that is particularly common in the Canadian Arctic and Alaska. It is absent from Australia which, when part of the east coast of Pangea, had a warm climate (Bambach et al. 1980).
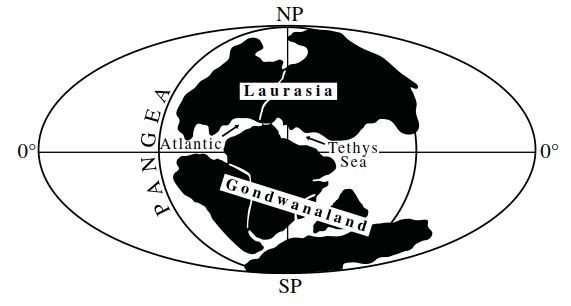
Figure 4.8. Map showing the break-up of Pangea in the late Triassic era (180 million years BP)
Similarly, Artemiopsis, a presumptive Laurasian cold-water genus, is restricted to cold northern regions of Eurasia and North America (e.g. the Northwest Territories of Canada). Conversely, Branchinella and Artemia, which are warm-water forms, are absent from modern-day Antarctica. The limited distribution of Artemia in Australia may be due, in part, to competition from Australia's native brine shrimp, Parartemia (Geddes 1980). The absence of strepto- cephalids from Australia and South America suggests that Streptocephalus (another presumptive Laurasian genus) colonized Africa after the breakup of Gondwanaland and has since undergone a major adaptive radiation there—though mostly in the temperate zone (Brtek 1974; Belk 1981). Streptocephalus sealii is the most widely distributed anostracan in the United States (27 out of 48 states). In contrast, 18 of the 43 species known from the United States have been reported from only a single state each (Jaas and Klausmeier 2000). Anostracan species richness, in general, is always highest in temperate parts of the globe. In an attempt to account for climate being an important controlling factor in their distribution, Belk (1981) compared the anostracan faunas of Arizona and South India, two climatically different regions. Arizona is a region in the temperate zone that experiences marked seasonal differences in climate and includes environments that range from lowland desert to high alpine.
The anostracan fauna is represented by 13 species from five genera (Artemia, Branchinecta, Eubranchipus, Streptocephalus, and Thamnocephalus). South India is a region in the tropical zone that experiences only slight seasonal changes in climate. The area of the Western Ghats mountains and to the west is humid and anostracans are not found there, but east of the mountains is a semi-arid area where temporary ponds are very common. Here the anostracan fauna is represented by only six species from three genera (Artemia, Branchinella, and Streptocephalus). The variation in thermal regime associated with refilling of temporary ponds is much less in southern India than in Arizona, and Belk maintained that because there are significant differences between the temperature requirements for egg hatching between species, a lower diversity is to be expected in the less variable environment. Southern Africa is particularly rich in anostracans, with 80% (38 species) being endemic. However, the fauna is under threat from agricultural practices, urbanization, pollution, and pesticides (Hamer and Brendonck 1997).
In the Sahel, Senegal, downwind drift from routine application of a variety of insecticides used to control Desert Locusts has been shown to be detrimental to anostracans and cladocerans living in local ponds (Lahr 1998). Vekhoff (1997) recorded a comparatively rich branchiopod fauna from the high arctic islands and archipelagoes of the Barents Region of Russia (four anostracans, one notostracan, and two con- chostracans). This high diversity was thought to be the result of the fauna comprising both widespread tundra species and species that had dispersed into the region from the nearby mainland. One species, Branchinecta paludosa, occurred in 45-50% of the crustacean communities, and reached its most northerly distribution (77°N) at Ivanov Bay.
There are also global and regional differences in the distribution of the Notostraca (tadpole shrimps). Rzoska (1984) stated that the genus Lepidurus is northern and may occur in Arctic conditions, whereas the genus Triops (—Apus) is confined to warmer waters, especially in arid regions and hot climates. In the United States, Triops longicaudatus seems to be confined to the drier western half of the country, and its range extends down into Mexico, the West Indies, the Galapagos Islands, Argentina, and the Hawaiian Islands. There is one record for Canada, in Alberta (Figure 4.9). Recent electrophoretic evidence, however, suggests that T. longicaudatus is actually a mixture of at least two reproductively isolated species (T. longicaudatus and T. newberryi)—the former typical of central, prairie pools, the latter more characteristic of large, southwestern, playa pools (Sassaman et al. 1997).
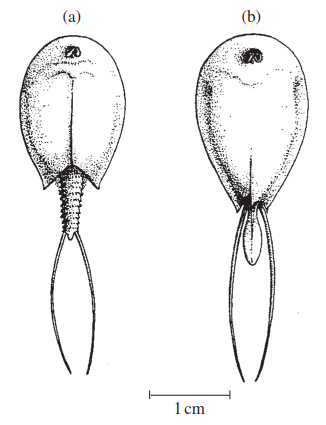
Figure 4.9. Notostraca: (a)—Lepidurus arcticus Pallas 1793; (b) —Lepidurus couesii Packard 1875
The genus Lepidurus is represented by several species in North America and these again occur chiefly in the western states but also in central and arctic Canada. Lepidurus arcticus, (Figure 4.10), for example, is found only in Alaska, the Northwest Territories and Labrador, plus in Greenland (Figure 4.9). Lepidurus couesi (Figure 4.10) is found in Montana, North Dakota, Oregon, Idaho, and Utah in the United States; Alberta, Saskatchewan, and Manitoba in Canada; and in Russia, Northern Siberia and Turkestan in the Palearctic (Linder 1959; National Museum of Canada records).
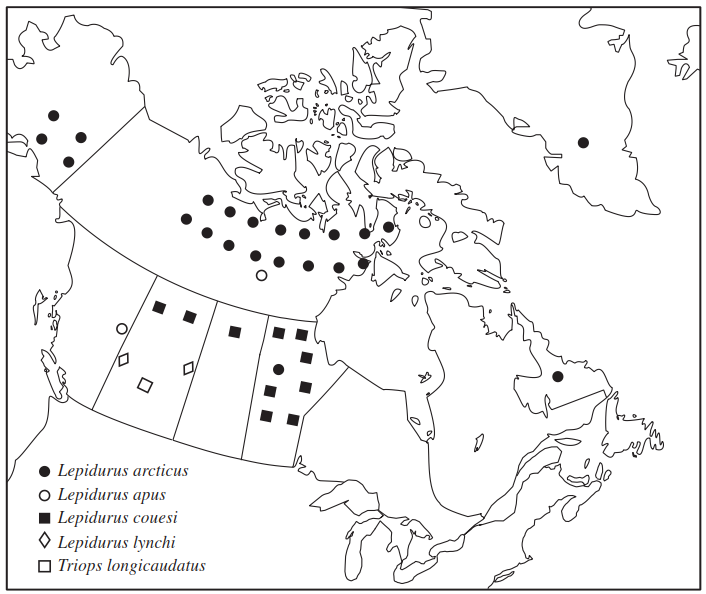
Figure 4.10. Records of anostracans from Canada, Alaska, and Greenland (dots represent single collections made in a particular province/state—they do not accurately depict where the collections were made. Based on records in the Canadian Museum of Nature, Ottawa, and in Linder 1959)
In Australia, both Lepidurus and Triops occur in virtually all states except Tasmania, where only Lepi- durus is known; only Triops has been found in the Northern Territories. However, Lepidurus is largely confined to the more temperate southeastern and southwestern corners of the continent, while Triops tends to be more common in the dry interior, although it also occurs on the coast in a few regions (Figure 4.11).
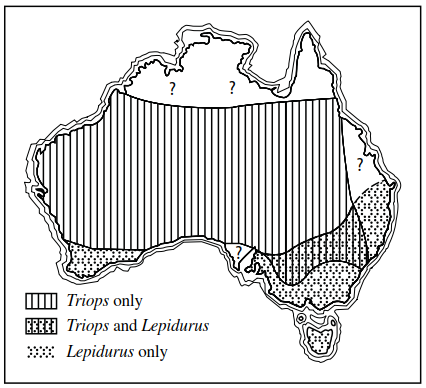
Figure 4.11. Geographical distribution of Triops and Lepidurus (Notostraca) in Australia (redrawn after W.D. Williams 1968)
These distribution patterns seem to be correlated with regional differences in climate particularly in terms of mean annual temperature and evaporation (Figure 4.12). Even though the ranges of these two genera overlap in the southeast, they are never found coexisting in the same body of water and this appears to be true for much of the rest of the world also (Williams 1968).
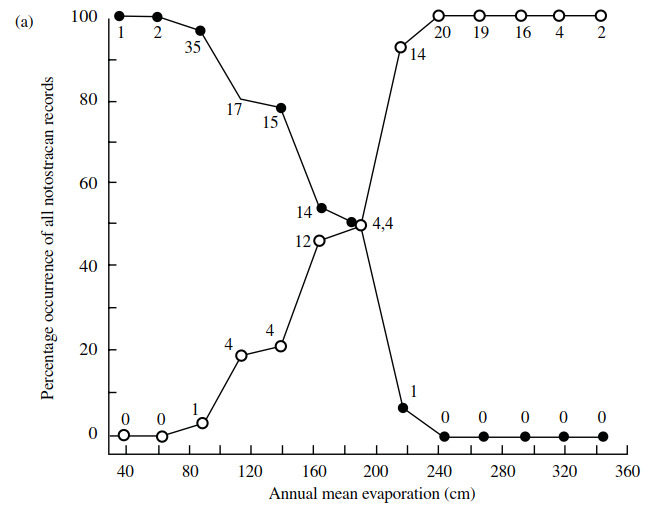
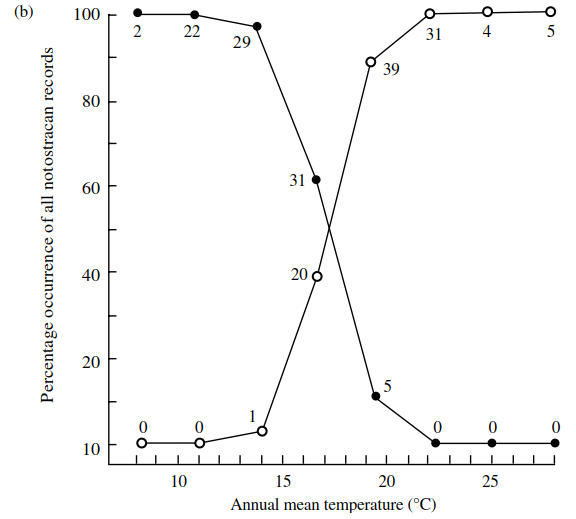
Figure 4.12. Distribution of Triops (open circles) and Lepidurus (closed circles) (Notostraca) with respect to: (a)—isoclines of annual mean evaporation; (b)—isotherms of annual mean temperature (numbers of records are given; redrawn after W.D. Williams 1968)
The Conchostraca (clam shrimps) have a wide geographic distribution primarily in temporary waters but also in the littoral zone of lakes. In general, they are found in warmer waters than most anostracans. In common with other large branchiopods, their preference for temporary waters is believed to be a stratagem for avoiding predatory fishes against which they seem relatively defenceless. Some species have very extensive distributions while others are known only from their type locality. For example, in North America, Lynceus brachyurus is found across the United States as well as in most Canadian provinces (except for the Maritimes), the Northwest Territories and Alaska (Figure 4.13).
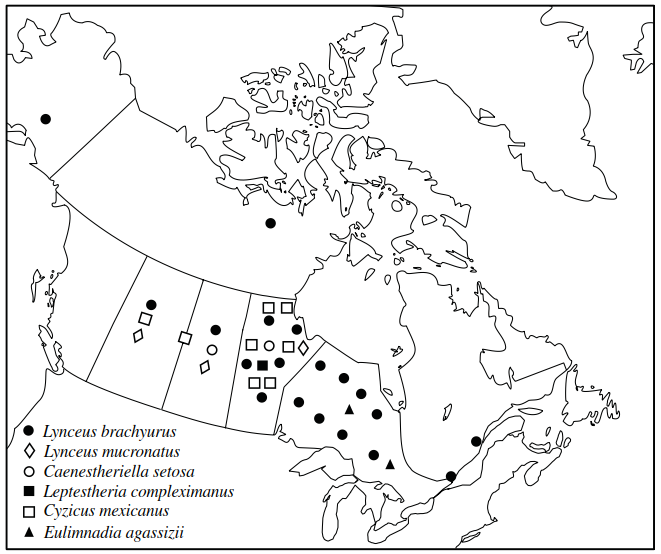
Figure 4.13. Records of conchostracans from Canada and Alaska (dots represent single collections made in a particular province/state—they do not accurately depict where the collections were made. Based on records in the Canadian Museum of Nature, Ottawa, and in Maddox 1959)
It is also found in Europe and Asia. Cyzicus mexicanus occurs across most of United States and Mexico, and also in Alberta and Manitoba (Mattox 1959; National Museum of Canada records). Some species of Eulimnadia, however, have very restricted distributions. Eulimnadia alineata and E. oryzae, for example, have been recorded only from rice fields at Stuttgart, Arkansas (Mattox 1959). In Colombia, Eulimnadia magdalenensis seems highly adapted to very short-hydroperiod ponds in arid regions, whereas E. colombiensis prefers longer- lasting ponds in cooler locations (Roessler 1995). Three species of Eulimnadia have also been recorded from Australia (from the Northern Territories, central and Western Australia, Queensland, and Tasmania). In total, there are some 23 species of Conchostraca in Australia which are all, with the exception of Cyclestheria hislopi, endemic (Williams 1980). Cy. hislopi has a circumtropical distribution and is one of the few conchostracans capable of existing in permanent waters. Here, it demonstrates effective anti-fish predator behaviours including: hiding among vegetation; production of a special, protective mucous capsule; and direct development, in which larvae leave the safety of the maternal carapace only when they are capable of secreting their own capsule (Roessler 1995).
Most members of the Malacostraca (Amphipoda, Isopoda, Mysidacea, and Decapoda) lack the ability to deal with desiccation and have no capability for diapause, adaptations that would suit them to life in temporary waters. There are, however, a limited number of exceptions. The amphipod Hyallela azteca, for example, is a widespread species in permanent waters in North America, but is occasionally found in intermittent ponds and streams where it appears to survive the summer in small, isolated pools. In the wetlands of western Canada, H. azteca along with Gammarus lacustris occur at such high densities that they influence the abundance and distribution of waterfowl, specifically the lesser scaup (Aythya affinis) (Lindeman and Clark 1999). Two species of Crangonyx, C. setodactylus and C. minor, are common in intermittent streams in eastern North America where they reproduce and grow during the lotic phase, but retreat into moist substrate or share the burrows of crayfishes as the hydroperiod ends (Williams and Hynes 1977). There are very few records of isopods from temporary waters. Apart from terrestrial forms, such as species of Oniscus that forage on bed detritus during the dry phase, only Caecidotea is recorded in the North American literature (Batzer et al. 1999). Paucity of peracarids in temporary waters is perhaps surprising given that many species occur in subterranean and interstitial habitats—there are around 116 species of amphipod that live in groundwaters in North America, alone (Covich and Thorp 1991).
In contrast, many decapod species are associated with temporary waters, in particular those belonging to the Astacidae (crayfishes) and Brachyura (crabs), and occasionally the Palaemonidae (shrimps). They are able to live in these habitats because of a range of adaptations that include: physiological adaptations—such as an ability to regulate their oxygen consumption (seen in some cambarid crayfishes); life cycle modifications— such as brooding of their young until an advanced stage; and behavioural adaptations—such as burrowing into the substratum to follow retreating groundwater tables. Burrowing behaviour in North American crayfishes has been categorized into: (1) primary burrowers (e.g. Fallicambarus devastator), which typically live most of their lives within complex tunnels that may be some distance from open water (Figures 4.14(a,d)); (2) secondary burrowers (e.g. F. hedgpethi), which also tend to live mostly within their burrows, but seek out open water during rain events (Figure 4.14(b)); and (3) tertiary burrowers (e.g. Procambarus acutus), which live in open water when it is available, but retreat into burrows at other times (e.g. to brood their eggs, to avoid desiccation, and to move below the frost line in winter; Figure 4.14(c,e)). Within their burrows, individuals do not always immerse themselves in water, as they have the ability to respire in humid air. Some non-burrowing species are also capable of digging simple shafts in bed materials to avoid drought (Hobbs 1991). During drought, a variety of other species are known to share the refugial water of crayfish burrows (see Chapter 5).

Figure 4.14. Different forms of North American crayfish burrows in relation to ground- and surface water levels (a and d—primary burrowers [see photo of Fallicambarus fodiens]; b—secondary burrowers; c and e—tertiary burrowers; redrawn after Hobbs 1981)
Crayfishes are known to have a significant impact on temporary water environments. For example, when Procambarus clarkii was introduced into a freshwater marsh in Spain, it altered the structure and function of the marsh community— primarily through impact on the pre-existing food web (Gutierrezyurrita et al. 1998).
Several species of true crab (Brachyura) live in brackish- or freshwater wetlands. Indeed, such habitats may have played a significant role in the evolution to terrestrial life seen in, for example, some of the grapsid crabs of Jamaica. There are nine endemic species of land crab on this island, some of which show active brood-care for their larva and juveniles, a phenomenon not known elsewhere in this group. These species demonstrate a range of dependence on water along with specific adaptations to a variety of habitats. The most specialized of these are Metopaulius depressus, which raises its larvae in water-filled bromeliad leaf axils, that it defends against predators, and Sesarma jarvisi, which breeds in empty snail shells that it keeps topped-up with rainwater (Schubart et al. 1998).
Microcrustaceans are represented in temporary waters by the smaller branchiopods (Cladocera), copepods, and ostracods. Cladocerans are typically found in lentic habitats, and many species have cosmopolitan distributions—although some of the latter may be confounded by taxonomic inadequacies. Many species appear to be capable of producing haemoglobin facultatively, when faced with low oxygen levels. Reproduction is par- thenogenetic and rapid (often just 1 or 2 days) for much of the hydroperiod, and the number of instars varies according to both genetic and environmental factors. Males appear close to the end of favourable habitat conditions (pre-winter, or pre-drought) and sexual, resting eggs are produced thereafter. These eggs are protected by an ephippium and can withstand freezing, drying, and dispersal to new habitats. Many cladocerans exhibit a wide tolerance of environmental conditions such as pH, temperature, and dissolved oxygen, yet others are restricted to very narrow ranges of water chemistry, especially dissolved calcium and salinity. In permanent waters, plank- tivorous fishes are believed to be influential in structuring cladoceran communities; in temporary waters they may be affected by invertebrate predators such as odonates and beetles.
Most of the 12,000 known species of copepod are marine. However, species belonging to the three main orders, Calanoida, Cyclopoida, and Harpacticoida have successfully colonized most freshwater habitats, in all climatic zones. Temporary waters in which they live include snow-melt water pools, desert pools, phytotelmata, damp moss, and wet leaf litter on forest floors and in compost bins. Reproduction is largely by sexual means (except in a few harpacticoids), with fertilized eggs being carried, and possibly still nourished, by the female for several days until their release. There then follow six naupliar stages, followed by six cope- podite stages, the last being the adult. Development time can take as little as 1 week, depending on habitat conditions. Diapause is a dominant feature in the life cycle of many species. Harpacticoids and calanoids are capable of producing two types of eggs: subitaneous eggs that hatch within a few days of being laid, and resting eggs capable of long periods of dormancy. Production of egg type seems to depend, at least partly, on environmental cues. Cyclopoids produce only subitaneous eggs, however individual animals may enter a resting stage as late copepodites. The latter may be either a simple developmental arrest, or full diapause (Williamson 1991). Harpacticoids are also capable of near-adult diapause under unfavourable conditions. More information on the adaptations of crustaceans, in general, to temporary waters will be given in Chapter 5.
Ostracods are very common in temporary waters but records often lack detail due to difficult taxonomy—although this is now improving. Fossil records show that the earliest freshwater forms lived in coal-forming swamps, ponds, and streams of early Pennsylvanian age (Benson 1961). Reproduction may be either sexual or asexual, and eggs may hatch soon after being laid, or may be staggered over time. Eight moult stages are passed through before the mature adult emerges. The eggs of freshwater ostracods have a structure (a double wall of chitin impregnated with calcium carbonate, with a fluid-filled space in between) that endows great resilience to environmental adversity, especially desiccation and freezing (Delorme 1991). Drought resistance was one of the criteria listed by Marmonier et al. (1994) as characterizing species found in temporary ponds on the floodplain of the Upper Rhone River—the others being: high reproductive rates, short life span, spherical shape, long swimming bristles, and low thigmotaxis. For North American temporary ponds, Delorme (1991) cited the following species association as being typical: Candona renoensis, Cypricercus deltoidea, Megalocypris alba, and Cyclocypris laevis. In some habitats, such as the semi-arid and arid regions of southern Africa, diversity and endemicity are remarkably high, comparable only to the ostracod faunas of very stable habitats, for example, ancient Lake Tanganyika (Martens 1996).
As shown in Chapter 3, from North American data (Table 3.2), the length of the hydroperiod seems to significantly influence both macro- and microcrustacean diversity (see also Ebert and Balko 1987), with the greatest number of species occurring in habitats that contain water for between 150 and 250 days in a year. Apart from the anos- tracan Eubranchipus and the conchostracan Lynceus, microcrustaceans (comprising three cladocerans— Daphnia, Scapholeberis, and Simocephalus, the cala- noid Leptodiaptomus, and several cyclopoid copepods) persisted in habitats containing water for less than 70 days. Further, the cyclopoids and Scapholeberis survived in hydroperiods of less than 10 days per year. In a study of English wetlands, Jeffries (2001) showed that, at the species level, the relationship between length of the dry period and the subsequent presence of microcrustaceans was negative for some but positive for others, probably contingent on the characteristics of individual ponds. Laird (1988) found that cladocerans (Chydorus sphaericus and Daphnia middendorffiana), an ostracod (Cyclocypris globosa), a cyclopoid (Acanthocyclops vernalis), and unidentified harpac- ticoids were the only crustaceans present in his northern Quebec snow-melt pool (hydroperiod more than 3 weeks). Additionally, a species of the cladoceran genus Moina, and two cyclopoids (Mesocyclops thermocyclopoides and Microcyclops varicans) were the only two crustaceans in his episodic puddle (hydroperiod less than 9 days) in Singapore. Mature specimens of Moina dubia were recorded within 2 days of refilling in a short-lived tropical rainpool in the Sudan (Rzoska 1961). Numerically, Lahr et al. (1999) found cladocerans to be the dominant taxon present early on in the 7-month hydroperiod of a temporary pond in the arid Sahel region of Senegal. Later, the ponds were dominated by 'more slowly establishing' crustaceans, particularly copepods, which then gave way to insects.
Date added: 2026-07-14; views: 3;