Insects and Arthropods in Temporary Waters: Ecology
A low-resolution view of the major groups of aquatic insects (including the Collembola) known to occur in temporary running waters is presented in Figure 4.15. The horizontal axis in this figure represents a relative scale of habitat permanence. Habitats such as stable, coldwater springs are placed at the high permanence end, while temporary streams are placed at the low permanence end. Extending this logic, episodic streams would be placed at a lower permanence level than intermittent streams. However, as there has been little distinction between these two latter habitat types in the past literature, the present analysis necessitates combining distributional records under the more general category of 'temporary' streams. In temporary lotic waters, there appear to be no records for the Neuroptera and Lepidoptera (although pyralid larvae have been collected from a stream in southern Australia; A.J. Boulton, Personal Communication), and the Plecoptera and Odonata are poorly represented. Particularly well-suited to temporary streams are the Ephemeroptera, Hemiptera, Coleoptera, Trichoptera, and Diptera. Higher resolution (Figure 4.15) shows that the Tipulidae and Chironomidae (Nematocera) dominate the temporary stream dipteran fauna (Williams and Feltmate 1992), whereas brachycerans are comparatively rare.
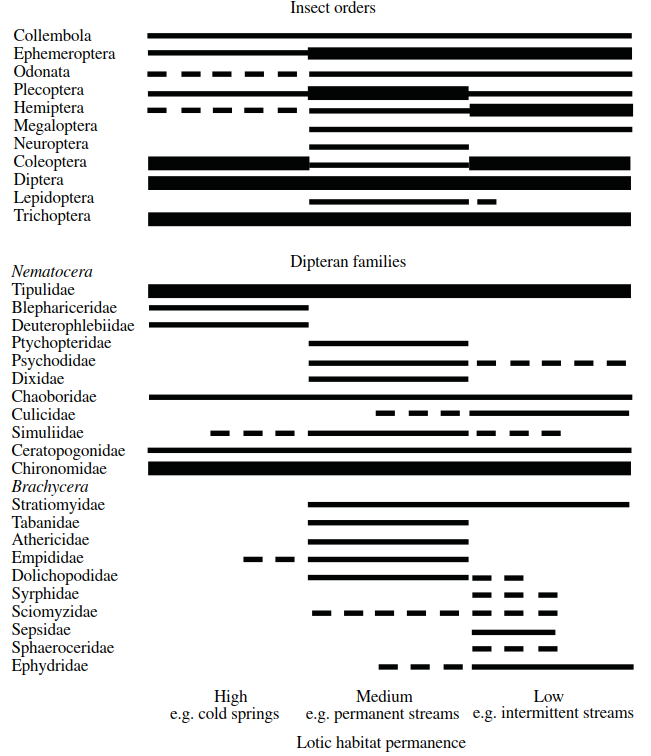
Figure 4.15. Overview of the relative importance of different aquatic insect orders and dipteran families across a gradient of running water habitat permanence (thick line indicates ‘well represented’, thin line indicates ‘moderately well represented’, broken line indicates ‘present but not significantly so’; information taken mainly from Merritt and Cummins 1984, and Williams and Feltmate 1992)
With the exception of mayflies (although a few species may attain large populations in vernal pools, Wiggins et al. 1980), all of the orders commonly found in temporary running waters also are common in temporary ponds (Figure 4.16). In addition, the odonates are well-represented. The most common nematoceran Diptera in temporary ponds are the Tipulidae, Culicidae, Ceratopogonidae, and Chironomidae, together with the brachyceran Ephydridae. Odonates and dipterans also are well-represented in highly specialized habitats such as phytotelmata (Corbet 1983; Beaver 1985).

Figure 4.16. Overview of the relative importance of different aquatic insect orders and dipteran families across a gradient of standing water habitat permanence (thick line indicates ‘well represented’, thin line indicates ‘moderately well represented’, broken line indicates ‘present but not significantly so’; information taken mainly from Merritt and Cummins 1984, and Williams and Feltmate 1992)
A number of observations can be made from these data. First, they indicate that, at the level of orders and families, temporary waters do not support diverse insect faunas as are found in permanent waters. Second, some very general inferences can be made about the adaptive traits and trade-offs demonstrated by different taxa. For example, it appears that Plecoptera seem less able to deal with standing water conditions than with drought, and survive drought only in running water habitats. Conversely, several dipteran families (e.g. Ptychopteridae, Tabanidae, and Empidi- dae) are capable of living in temporary standing waters but not apparently in temporary running waters (although there are records of empidids from a semi-permanent spring in Germany and from two temporary streams in Australia; F.O. Gathmann, Personal Communication, and Boulton and Lake 1992a, respectively). Despite their considerable powers of flight, Trichoptera and aquatic Hemiptera and Coleoptera do not colonize phytotelmata significantly. Neuropterans seem not to have developed the capability of living in temporary habitats even though they are found in a range of permanent waters.
Although some of these conclusions are tentative, due to inadequate study of temporary waters, they nevertheless point to some interesting avenues for future research into evolutionary trends in habitat selection among aquatic insects. Particularly useful would be determination of why the Plecoptera, Odonata, and brachyceran Diptera are poorly represented in temporary streams, yet the Tipulidae and Chironomidae are successful; why mayflies are not widespread in temporary ponds despite being occasionally abundant locally; what mechanisms, physiological or otherwise, result in decrease in species diversity with decrease in the length of the aquatic stage of the habitat; and why the Trichoptera, Hemiptera, and Coleoptera are poorly represented in phytotelmata. In addition, there needs to be more extensive surveying of habitats in order to confirm the global nature of temporary water communities and to extend/ confirm the known ranges of taxa along the permanent-temporary water habitat gradient.
The relationship between a habitat permanence gradient and community structure can best be examined when a dataset contains a high level of taxonomic resolution. Some examples of this relationship, for insects, have already been given in Chapter 3 (e.g. Table 3.1). Another, intruiging insect dataset is provided by Wissinger et al. (1999), who compared the insect species found in 41 subalpine wetland basins in Colorado. These basins were divided into three groups, based on their number of open-water days: 40-64; 74-116; and greater than 130 (Table 4.4). The taxon counts were 10, 26, and 50, respectively. Clearly, numerically, these data support the trend of decreasing species richness with decreasing length of the hydroperiod. However, examination of the taxa in the >130-day hydroperiod group, which was designated 'permanent' by the authors, reveals quite a number of taxa frequently seen in temporary waters, such as the hemipterans; the beetle genera Helophorus, Agabus, Acilius, Dytiscus, Rhantus; the dipterans Bezzia and Stratiomyia, and several chir- onomid genera. The interesting point that emerges about the >130-day hydroperiod group is that although none of these basins has dried out completely in the past 50 years, they are only free of ice and snow cover for about 4 months of each year, and most are frozen to within just a few centimetres of their beds. It could be argued, therefore, that they do not truly represent permanent water habitats, and that their short window of opportunity for colonization attracts pioneering taxa, resulting in a community with distinct temporary water elements.

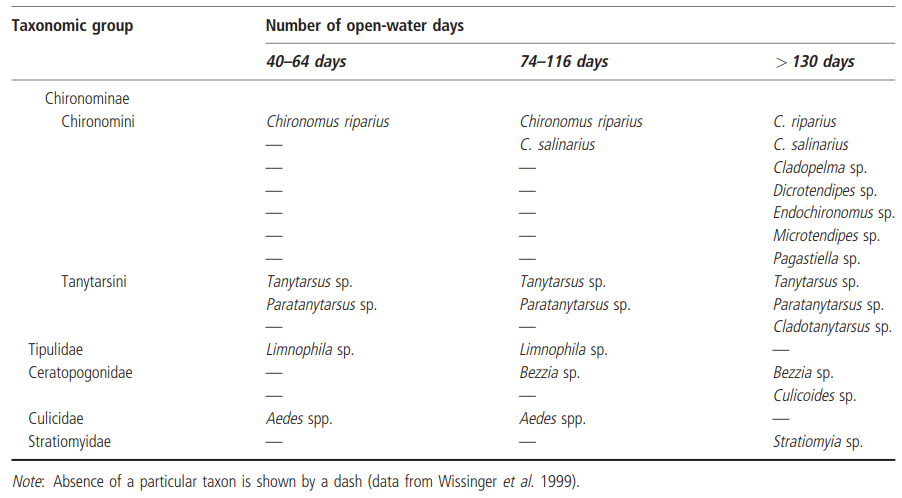
Table 4.4. Comparison of the dominant insect taxa in 41 subalpine wetland basins in Colorado with different degrees of permanence (shown as the number of open-water days in the year)
In addition to the largely aquatic taxa described above, there are a number of other insects associated with temporary waters, especially at the end of the hydroperiod, when there is decaying aquatic vegetation and moribund aquatic prey to be harvested. Little attention has been paid to these, and other arthropod taxa (see below), thus their roles in temporary water ecosystems are poorly understood, but perhaps important. Groups of note are the grylloblattids (rock crawlers), ants, wasps, and various terrestrial beetles.
Lude et al. (1999) found that at least nine species of ant were able to survive frequent inundation on Alpine floodplains. In particular, Formica selysi regularly colonized relatively young, unvegetated gravel islands and bars, and survived flooding by forming swimming rafts when its nest entrances were compromised. Each raft consisted of several dozen workers, a queen, and brood, and remained intact until it reached the shoreline. Clearly, this ant species found suitable foraging opportunities on these periodically flooded areas. Other hyme- nopterans associated with seasonally flooded sites include wasps and bees. For example, Visscher et al. (1994) recorded the ground-nesting bee Cal- liopsis pugionis emerging from sites that had been underwater for more than 3 months in the floodplain of the San Jacinto River. The authors suggested that the flooding regime influenced the sex ratios of bees as they emerged from diapause.
Carabid beetles are known to be commonly associated with damp environments, including wetlands, floodplains, etc. Basta (1998) recorded 94 species from a single marsh in the Czech Republic, where population densities of some of the most abundant species seemed tied to fluctuations in water level. Carabids were also identified as a major component of the riparian fauna on four alpine floodplains in Bavaria, where most of their prey were aquatic species. In particular, on the Isar floodplain, river-derived invertebrates represented 89% of carabid prey, primarily emerging chironomids (fed upon by small species of Bembidion) and stoneflies (favoured by Nebria pici- cornis) (Hering and Plachter 1997). Juliano (1985) found a variety of carabids of the genus Brachinus associated with different pond types in Arizona. Brachinus lateralis dominated the margins of more permanent ponds, whereas high-elevation temporary ponds were dominated by B. mexicanus. Brachinus javalinopsis and B. lateralis co-dominated the margins of a low-elevation temporary pond, but only B. mexicanus was found in dry pond basins.
All three species were believed to share at least two potentially limiting resources: food for the adults (carrion and other arthropods), and water beetle pupae (required as hosts for their ectoparasitoid larvae). Lott (2001) reported that temporary ponds in lowland England support a rich beetle fauna, comprising chiefly carabids and staphylinids. Some of the species are those associated with the margins of larger, pemanent water bodies and river floodplains. Assemblages differ among ponds, and at least part of this can be attributed to bed substrate composition (e.g. mineral versus peat). A number of interesting adaptations to being occasionally submerged by water have been observed (e.g. retreating to air pockets in the bed litter, retreating up the bank prior to a flood advance, or skimming over the water surface after secretion of a water repellant), however, the ecology and behaviour of most species are largely unknown.
Arachnids and other arthropods The arachnids most commonly found associated with temporary waters can be divided into three informal groups: water mites, which are primarily associated with the hydroperiod; soil mites, which are more likely to be associated with the moist sediments of the basin after the water has evaporated; and spiders, which, apart from some aquatic and semi-aquatic forms, live in the riparian zone, but also may move onto the drying bed to scavenge.
Water mites are considered as those belonging to five somewhat unrelated subgroups of small arachnids. The Hydrachnida is the most familiar, but some forms belonging to the Oribatida, Halacaridae, Mesostigmata, and Acaridida (the latter three being of minor importance) have invaded freshwaters, giving rise to species that are now fully adapted to an aquatic existence. Water mites may be extremely abundant in ponds and the littoral zones of shallow lakes, for example, 2,000 individuals m~2, representing as many as 75 species. Many mites represent important micropredators in temporary waters, and some have coevolved with major insect groups (especially the Diptera), both parasitizing their bodies and using their adult stages as vessels of dispersal to new habitats (Smith and Cook 1991). In North America, families/subfamilies of hydrachnids commonly found in temporary waters include the: Hydrachnidae, Eylaidae, Piersigiinae, Hydry- phantinae, Thyadinae, Tiphyinae, Pioninae, and Arrenurinae. In general, hydrachnids are able to survive in temporary waters by one of two means: physiological endurance, or avoidance—for example, larvae of the Eylaidae and Hydrachnidae can remain attached to their adult insect hosts for the entire duration of the dry phase (Wiggins et al. 1980; Smith and Cook 1991).
The ecology of oribatid mites is not well known, partially due to problematic taxonomy. However, they are extremely abundant in most forested ecosystems and often comprise around 50% of the total microarthropod fauna. As such, oribatids are believed to play important roles in the decomposition of organic materials, modification of the physical and chemical textures of soil, the cycling of nutrients, and the conservation of healthy soil environments (Wallwork 1983). Habitat features thought to influence their population biology include habitat complexity and soil micropore size, soil humidity and organic content, soil temperature, surface vegetation, precipitation, and the activity of other soil microfauna (Wauthy et al. 1989). Because forest soils are often contiguous with temporary pond and stream beds, it is likely that oribatid mites contribute to biological processes (largely unknown) in these sediments. That there are characteristic assemblages of these mites in temporary waters is illustrated by the fact that oribatids have been found useful in distinguishing between different stages of degradation of freshwater mires in the area of Berlin (Kehl 1997). In coastal freshwater habitats in Antarctica, two species of terrestrial oribatids, Edwardzetes elongatus and Trimaloconothrus flagelliformis, have become adapted to survive prolonged submergence on aquatic mosses—apparently in response to niches unoccupied by truly aquatic forms (Pugh 1996).
There is only one species of truly aquatic spider. Argyroneta aquatica (Agelenidae), found in ponds of central and northern Europe, builds an underwater air-store from silk fashioned in the form of an inverted vase. It supplies this store with air brought from the water surface, and uses it as a medium in which to externally digest prey captured under water. Many other spiders, however, frequent the margins of both permanent and temporary ponds and streams. All members of the Pisauridae, for example, depend on being close to water, and some, such as Dolomedes dive while hunting for tadpoles, aquatic insects, and small fishes. There is a rich diversity of pisaurids in Australia, for example, including: Dendrolycosa (which also extends from New Guinea to India), Dolomedes (cosmopolitan), Hygropoda (also in southeast Asia, Venezuela, New Guinea, Madagascar, and central Africa), Perenthis (also in India, Burma, Papua New Guinea, and Japan), and the endemics Inola and Megalodolomedes. (Main et al. 1985). Another family of common riparian spider is the Tetragnathidae—the long-jawed orb weavers. The genus Tetragnatha has a wide global distribution, with T. versicolor and T. elongata being circumboreal. Williams et al. (1995) calculated that, as a minimum estimate, individuals of these two species captured 0.2% of the total number of insects (particularly chironomids and mayflies) emerging from a small river in Canada. Tetra- gnathid webs are common on any vegetation overhanging temporary ponds and streams, where their prey also includes emerging mosquitoes and other long-legged dipterans. The diversity of spiders associated with temporary waters can be very high, for example, van Helsdingen (1996) recorded a total of 63 species from two Irish floodplains.
As introduced above, a number of other, noninsect arthropods are associated with the end of the hydroperiod. Whereas zoogeography will dictate local composition, this 'clean-up crew' may include millipedes, centipedes, symphylans, pauropods, pseudoscorpions, harvestmen (Opi- liones), diplurans, bristletails (Archaeognatha), and silverfishes (Thysanura).
Date added: 2026-07-14; views: 3;