Biological Factors Shaping Temporary Freshwater Biotas
Brief reference, above, to the role of organisms in basin substrate processing and nutrient release brings us to consider some of the biological factors that influence the biotas of temporary freshwaters— summarized in Figure 3.8. Whereas it is not difficult to identify the influence of single biological factors, assessing their relative importance and linkage is less easy owing to lack of research specifically on temporary waters. Strong biological features include the succession of species so characteristic of temporary waters (e.g. Williams 1983; Jeffries 1994), seasonal influx of aerial colonizers (Fernando 1958; Nilsson and Svensson 1994), and temporal changes in the food base—as surplus bed nutrients from the dry phase are used up, aquatic-phase fungal and other decomposers continue to process allochthonous bed materials, and phyto- and zooplankton become established (Barlocher et al. 1978; Wiggins et al. 1980; Maher and Carpenter 1984).
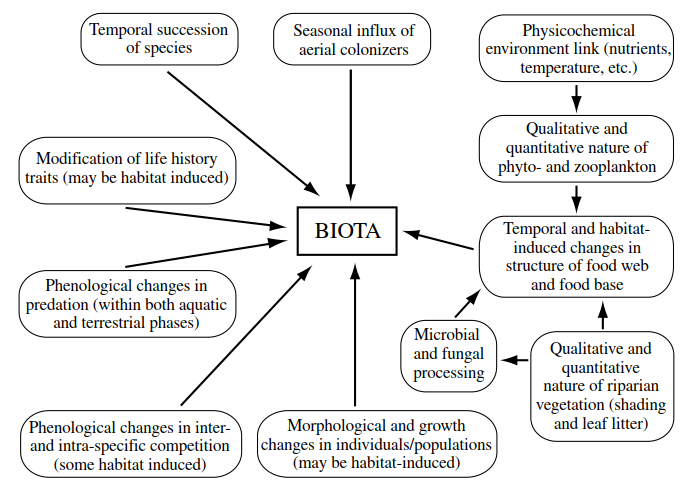
Figure 3.8. Summary of biological factors that influence the biotas of temporary fresh waters (from Williams 1996; see Figure 3.1 for explanation of arrows; note that interaction lines between factors have not been added due to uncertainty stemming from the lack of research on these factors specifically in temporary waters)
As a result of changes in the food base, together with variation in the numbers of predators (both as aquatic species succeed each other and as terrestrial ones invade the shrinking habitat), the structure of the food web must be subject to change (see later discussion). So, too, will the extent and intensity of inter- and intraspecific competition (Morin et al. 1988; Johansson 1993), although it has been argued that, in general, biotic interactions may contribute relatively little to community structure given the demanding requirements of the physicochemical environment (Poff and Ward 1989). In addition, a number of changes affect individual insects, such as modification of growth, morphology, and life history (Harrison 1980; Landin 1980; Juliano and Stoffregen 1994), that may be classed as genotype- environment interactions. More detailed coverage of these largely population- and community-level factors is left until Chapters 5 and 6.
A biological aspect that has received relatively little study, apart from the shading effect mentioned above, is the link between forested temporary waters and the surrounding vegetation. While plant biomass is known to lie at the base of such communities, the qualitative and quantitative influence of leaf-litter input is poorly known, relative, for example, to permanent waters (e.g. Hynes 1975; Lake 1995; Webster et al. 1995).
When a winter-frozen, intermittent pond begins to fill in the spring ('vernal' pond), degradation of both autochthonous (basin grasses and weeds) and allochthonous (riparian trees) leaf-litter begins; in the case of 'autumnal' ponds, this process may have begun, briefly, before winter. Degradation processes in intermittent waters can be inferred using flooded riparian woodlands as models. Merritt and Lawson (1992) identified various abiotic and biotic factors in the floodplain environment that influence litter processing (Figure 3.9), and have emphasized the importance of leaf 'conditioning'. Initially, autumn-shed leaves that have accumulated in the pond basin are subject to physical fragmentation via wet-dry rain cycles, winter freeze-thaw cycles, and wind.
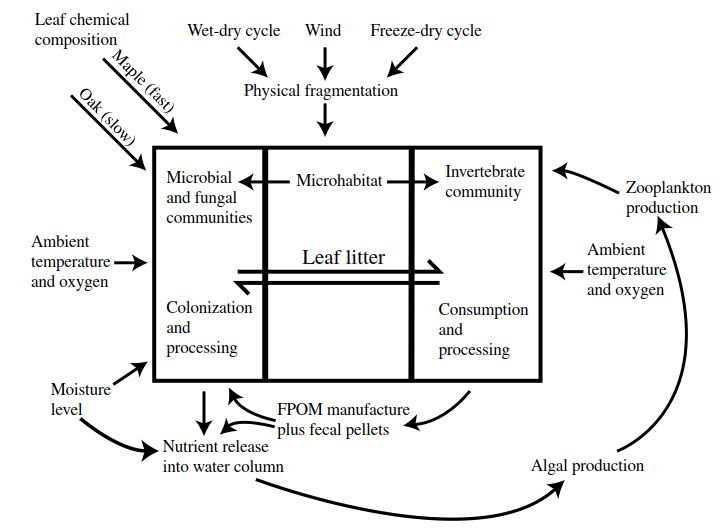
Figure 3.9. Schematic of the main abiotic and biotic factors associated with leaf-litter processing in forested temporary waters (based on Merritt and Lawson 1992)
Moisture levels and ambient temperature and oxygen levels are important as the basin begins to fill with water, and microbes and fungi colonize and begin to degrade the leaf pieces. At this stage, soluble organic compounds (e.g. sugars, polyphenols, and amino acids) are leached into the surrounding water (Suberkropp and Klug 1976). Leaf chemical composition is important in controlling the rate of these various processes, as some species (e.g. oak) are more resistant to conditioning than others (e.g. maple) (Kaushik and Hynes 1968). Release of leachates into the water column, together with other nutrients released by fungal and bacterial activity, contribute to an increase in benthic and planktonic algae, which in turn promotes the development of zooplankton populations. Leaf conditioning directly stimulates the growth of benthic invertebrate species, as aquatic hyphomycetes are know to be an important food source for many benthic invertebrates (Kaushik and Hynes 1971). As invertebrates consume and process leaf fragments, they manufacture FPOM (crumbs) and fecal pellets that are cycled back into the food web for consumption by others—frequently after recolonization by fungi and bacteria. In addition, whole leaves and fragments serve as microhabitats for pond inhabitants (e.g. as a physical substrate for microorganisms, and as shelter and attachment sites for invertebrates).
Date added: 2026-07-14; views: 6;