Prokaryotes in Temporary Freshwaters: Ecology and Adaptations
Introduction. The global temporary water biota is best known in terms of its higher plants and metazoans— although current studies are beginning to catalogue and understand the roles of bacteria, protoctists, and even fungi. However, it is the insects and crustaceans that dominate the fauna. The latter are well represented by micro-and macro-forms from the following major taxonomic groups: Branchiopoda, Ostracoda, Copepoda, Decapoda, Peracarida, and many rare species are present. Among the insects, it is the Hemiptera, Coleoptera, Trichoptera, and Chironomidae (Diptera) that are best represented. Aquatic mites (Hydracarina) are also common. Although typically absent from standing temporary waters, several fish species migrate in and out of temporary streams, rivers, and floodplains. Several types of amphibian live in temporary waters, some of them very rare, and temporary waters provide important feeding areas for migratory birds.
Tropical waters are more likely to have reptiles and mammals associated with them. This chapter will present summaries of each taxonomic group, providing examples of habitat-specific requirements and adaptations. The dearth of studies on the 'minor' taxa of temporary waters makes it difficult to discuss these groups in any other context than on the local scale. Case history studies of predominant temporary water types will be highlighted, ranging from the very large (floodplains) to the very small (rain-filled rockpools). Commonality in community composition, structure, and function will be sought. Whether any of the environmental influences discussed in Chapter 3 actually constrain the biota will be discussed.
Taxa. Prokaryotes.There are few inventories of named bacteria from temporary waters. Table 4.1 compares those found in an intermittent, snow-melt pool in northern Quebec with those in an episodic puddle in Singapore (Laird 1988). Although it is difficult to draw many conclusions from such a limited dataset, there are some similarities in community composition, despite the huge geographical separation and differences in predictability and lengths of their respective hydroperiods (the pool more than 3 weeks, the puddle no longer than 9 days).

Table 4.1. Comparison of the prokaryotes found in an intermittent, snow-melt pool in northern Quebec with those in an episodic puddle in Singapore
Felton et al. (1967) found nine physiological groups of bacteria in a temporary pond in Louisiana, USA. Sulphur-, ammonia-, and nitrite-oxidizing autotrophs were notably absent. It was felt that bacteria did not make a major contribution to the pond ecosystem as primary producers. However, counts of aerobic nitrogen-fixers and urea-using forms ranged from 106 to 108 per gram of bottom mud, suggesting a significant role in the nitrogen cycle in the pond. Bacteria capable of decomposing cellulose were found both in the water column and in the mud. Mud samples taken in the spring contained fewer cellulose-decomposers than samples taken in the autumn when, presumably, terrestrial plant material growing in the basin during the summer dry period would be starting to die and decay. Heterotrophs (those species requiring a supply of organic material from their environment) were the largest physiological group found in the pond, with anaerobes being more abundant than aerobes. Counts of both types decreased as the pond dried up but later increased after the basin was dry. Heat-shock experiments revealed that about 1.5% of the aerobic heterotroph population consisted of heat-resistant spores.
The authors concluded that the bacteria functioned as decomposers and transformers in the nitrogen, carbon, and energy cycles of the pond as well as acting as a source of nutrients and as a primary source of food for protoctists and plankton. Walker et al. (1991) have further highlighted the importance of bacteria as a direct food source for mosquito larvae. In field and laboratory microcosms, they were able to show that feeding by Aedes triseriatus reduced microbial densities, and that selective removal of larvae from tree holes and tyres was followed by rapid increases in microbial densities (Morgan and Merritt 1992). Kaufman et al. (1999) have shown that predation by Ae. triseriatus may also cause a shift in bacterial community composition in container habitats, from Pseudo- monaceae to Enterobacteriaceae. Larvae additionally affected microbial nitrogen metabolism, either through grazing on the bacterial taxa responsible for nitrification-denitrification, or through modification of the physicochemistry of the water.
Ectoenzymatic activities of cyanobacteria in the biofilm on Mediterranean stream substrates were found to resume significantly after only 2 h of re-immersion after drought. Romani and Sabater (1997) suggested that such rapid recovery was due to the stromatolitic (organosedimentary) structure of the biofilm, which acted as an organic matter reserve, alongside rapid rehydration permitted by the cell sheaths. This implies that such biofilms are capable of exploiting even short rainfall events during dry summers. The rapid re-establishment of the biofilm when streams resume flow ensures an almost immediate supply of food for grazers. Castillo (2000) has shown that populations and productivity of bacteria in neotropical floodplain lakes reach their highest values during low water, when they significantly exceed those of bacteria in the adjacent river water. In contrast, in blackwater lakes of Central Amazonia, bacterial abundance peaked during rising water—primarily due to input of allochthonous bacteria from the river at the beginning of the rainy season (Rai and Hill 1984).
Bacteria are well known for their tolerance extremes. In Don Juan Pond in the Antarctic, for example, where the freezing point of water is —40° C, bacteria have been observed growing in water temperatures ranging from —3 to —24°C (Hinton 1968). Many are also highly resistant to drying: of 2,724 bacterial strains tested, 83% of cultures dried in a vacuum for periods of up to 14 years, proved to be still viable (Rhodes 1950).
In general, functional studies of bacteria have been largely laboratory- or microcosm-based, and those that have been field-oriented have tended to concentrate on either purely aquatic ecosystems or terrestrial ones, although past methodologies have limited progress. Bacterial studies in habitats that experience a transition (e.g. from aquatic to terrestrial), such as temporary waters are rare, although rice fields are an exception (Conrad 1993). Aragno and Ulehlova (1997) have pointed out that transient ecosystems are likely to provide a multiplicity of ecological niches for a range of functional groups of bacteria, and are ripe for investigation with modern techniques. One such approach is terminal restriction fragment length polymorphism (T-RFLP), a phylogenetic method for the rapid analysis of complex microbial communities that has recently been applied to a freshwater ecotone (Sliva and Williams 2005).
A strictly numerical analysis of total bacteria in two adjacent intermittent ponds in Ontario has shown substantial differences both between ponds and within ponds in successive years. Mean densities in Pond I varied almost two orders of magnitude in the second year of the study, whereas in the same year, variation in Pond II was the least (Figure 4.1). Examination of environmental variables indicated that few were correlated with bacterial abundance in Pond I, but that the latter was strongly, and positively, correlated with turbidity in Pond II.
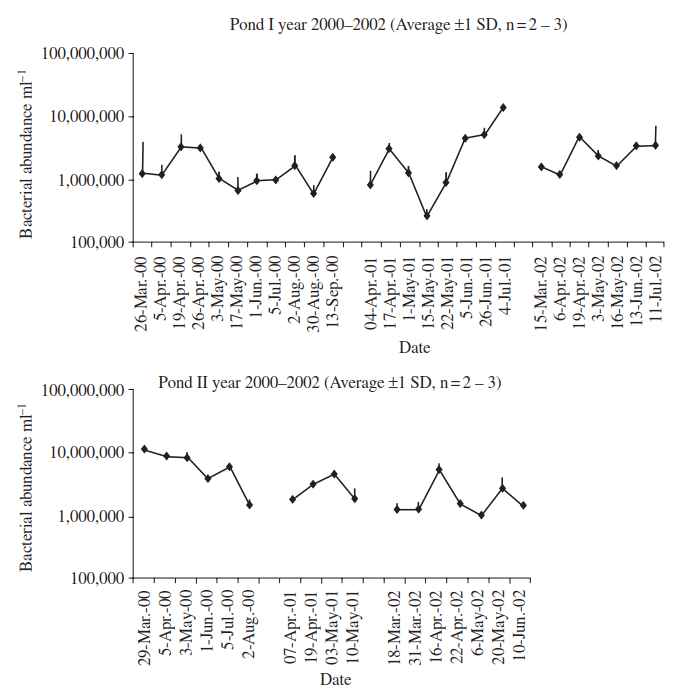
Figure 4.1. Comparison of the mean abundances (±1 SD) of bacteria in two intermittent ponds in southern Ontario throughout their entire hydroperiods, and over three years
A final aspect of temporary water bacteria worth mentioning is their role as pathogens. Again, the data are limited, but a vibrio bacterium has been found infecting the larvae of several container-inhabiting mosquito species in Florida (Fukuda et al. 1997). In addition, a baculovirus has been recorded from populations of the mosquito Wyeomyia smithii living in the pitcher plant Sarracenia purpurea, in Massachusetts (Hall and Fish 1974).
Date added: 2026-07-14; views: 6;