Substrate and Light Regulation in Temporary Waters
In both lotic and lentic temporary waterbodies, the size and type of bed substrate particles are likely to exert influences ranging from suitability as attachment sites during the wet phase to protection from desiccation during drought (Boulton 1989). As an example of the latter, Tabacchi et al. (1993) demonstrated that nymphs of the mayfly Thraulus bellus seek out the water-filled substrate interstices of temporary floodplain ponds as a refuge from fluctuating water levels. They also suggested that this behaviour may prevent overlap with populations of two other mayfly species, Caenis horaria, which lives in silt, and Cloeon gr. simile, which lives among macrophytes. Substrate composition also may be important to species survival. For example, the organic cases of larvae of the caddisfly Limnephilus coenosus proved to be better at holding water, and hence reducing mortality, in drying pools than the mineral cases of L. vittatus (Zamoramunoz and Svensson 1996).
In Chapter 2, it was described how various physical and chemical properties of vernal pool soils control the manner in which water drains from and returns to the basin (Weitkamp et al. 1996). Although, intuitively, the presence of a wetted substrate both prior to and at the end of the true hydroperiod would seem beneficial to the survival of early and late colonizers, little empirical evidence is available to support this. There is evidence, however, that soil type in flooded meadows influences microbial activity. Alternate flooding and draining has been shown to enhance nitrogen removal through stimulation of both nitrification and denitrification (Busnardo et al. 1992), with sandy soil removing more nitrogen than peaty soil. Infiltrating water may increase leakage of dissolved forms of both organic and inorganic nitrogen from the soil. Further, this water will also bring electron acceptors (nitrate-N, and dissolved oxygen) into the soil to promote mineralization (Davidsson and Leonardson 1998). Soil moisture and organic content in woodland floodplains has been shown to influence the abundance and distribution of larval tipulids (Merritt and Lawson 1981).
Apart from the role of microbes, there is a huge gap in our knowledge of other biological processes that must be presumed to take place in the wetted phases of temporary water basins. It is known that, in soils in general, besides bacteria, protoctists, and fungi, a number of metazoan groups are crucial to organic matter processing, particularly nematodes, earthworms, and arthropods. Among the latter, millipedes, insects, and a huge diversity of mites shred dead plant material, which begins the cycling of carbon, nitrogen and other nutrients by making available a large surface area for microorganism colonization.
These biological communities tend to be concentrated in the top several centimetres of substrate and around growing plant roots—as the rhizosphere attracts bacteria to sloughed-off plant cells and plant-released sugars and proteins. Fungal hyphae are common decomposers of plant debris and, unlike bacteria, can transport (via their spreading hyphae) nitrogen from the underlying soil to the litter layer. Fungi are also important as they are often the only organisms able to secrete enzymes capable of degrading complex compounds. Although nutrient release, which can benefit both terrestrial plants during the wetted phases and aquatic species when the water returns, is an important process, so also is the production of humus.
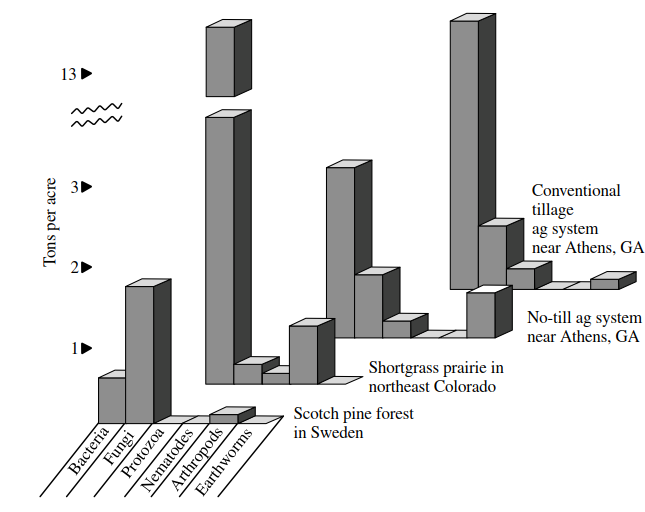
Figure 3.7. Data indicating that different soil types support different proportions of litter processing organisms (redrawn after Tugel and Lewandowski 1999)
Humus, or humidified organic matter, is what remains because it is not readily decomposed (it may be chemically too complex to be used by soil organisms, or it may be physically protected inside aggregates). Nevertheless, it is important in binding tiny soil aggregates and plays an essential role in improving the water and nutrient-holding capacities of the substrate. Different soil types support different proportions of litter-processing taxa (Tugel and Lewandowski 1999; Figure 3.7), which may well influence the organic matter breakdown rates and efficiencies in different temporary water basins, and hence perhaps the community composition and productivity of the aquatic phase. Further information on the organic component of bed substrates is given in Section 3.8.
Light. Light is of course an important factor in regulating primary production. Plant biomass lies at the base of most temporary water food webs, either as autochthonous algal and macrophyte tissue, or as the breakdown products of riparian leaf litter. Shoreline vegetation also may be influential in terms of shading, as this has been shown to contribute to the structure of the invertebrate communities of intermittent streams in Swedish meadows (J. Herrmann, personal communication). Photoperiod is a key aspect of light that is crucial to the survival of many temporary water species, as it provides essential cues that regulate the timing of life cycles, emergence from, and entry into, diapause, flight periodicities, and colonization dynamics. Responsiveness to photoperiod may vary with latitude, as has been shown for the pitcher-plant mosquito Wyeomia smithii (Wegis et al. 1997). More specific aspects of photoperiod will be covered in Chapter 5.
Date added: 2026-07-14; views: 5;