Temporary Pond Algae and Protozoans
Protoctists.Membership of this group follows that proposed by Margulis and Schwartz (1988), and includes both those phyla known as protozoans, and those grouped as algae. Some information on cyanobacteria is also included here, as the latter are often still included in general studies of algal communities.
Algae. Sheath and Hellebust (1978) studied the various algal communities in an arctic tundra pond and also reviewed the literature on these very abundant habitats. Tundra ponds develop where vegetation has been injured resulting in an increased thawing of permafrost, where there is a loss of volume as ice is melted, and where subsidence of sediments occurs. These ponds held water during the brief summer (June to August) but froze solid under 2 m of ice each winter. The plankton community showed two peaks in biomass and primary production during the short ice-free season. Only one peak in biomass was recorded for the periphyton community but primary productivity showed one or two peaks. Laird (1988) recorded a total of 114 species of diatom and 24 species of green alga from his snowmelt pond in northern Quebec.
Antarctic streams, which periodically lose their free water by virtue of freezing solid in winter, have been shown to support rich epilithic communities (87 taxa; Pizarro et al. 1996) of high biomass (>20 mg chlorophyll a cm~2, or >20 mgCcm~2), yet production rates are low (Vincent and Howard- Williams 1986). Nutrient supply and light were not thought to be limiting factors in these streams but water temperatures seldom rose above 5.0°C and, more often, lay between 0 and 2.0°C. Metabolism in these algae thus occurs at a low rate and the high biomass observed represents accumulation over several seasons of growth (as theoretical turnover times were of the order of several hundred days yet each annual growing season lasted less than 80 days). Despite this, the overwintering community retained a high metabolic capacity and responded rapidly to hydration at the beginning of summer.
Figure 4.2 shows that photosynthetic capacity of the cyanobacterium-dominated epili- thon rose as a log function of time over the first 6 h, and then at a faster rate over the subsequent two days. Thus, simple rehydration allowed immediate resumption of some photosynthesis but full recovery necessitated longer-term biosynthesis and repair. Vincent and Howard-Williams (1986) likened this resurrection to the response of desert plants in warmer regions, each community inhabiting a seasonally arid environment. They pointed out, however, that in addition to loss of water, Antarctic epilithon must contend with continuous darkness in winter and a harsh freeze-thaw cycle. Physiological resilience to freezing must therefore be an essential property of the microflora in the Antarctic. Fumanti et al. (1995) found a similarly rich microflora in Lake Gondwana, Northern Victoria Land, Antarctica. Although the phytoplankton was species poor (5 taxa), there was a rich benthic community in the form of shoreline mats comprising 34 taxa (8 species of green algae, 7 of diatoms, and 19 of cyanobacteria). Phormidium frigidium predominated in the lower mat layer, whereas the upper mat layer was dominated by Pleurococcus antarcticus (a green alga) alongside several other taxa all of which were characterized by gelatinous sheaths.
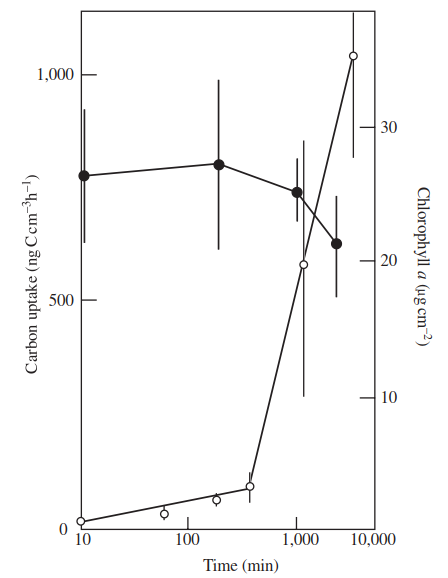
Figure 4.2. Photosynthetic recovery by the Phormidium epilithon as a log function of hydration time (closed circles represent chlorophyll a content; open circles represent photosynthesis; each point is the mean of three samples ±2 SE; redrawn from Vincent and Howard-Williams 1986)
In many types of temporary waters, the formation of algal mats, especially those formed from the drying and felted remains of filamentous species, can be crucial to the survival of other organisms that may take refuge under them during drought. Reynolds (1983) has recorded sheets of this 'algal paper' as large as 50 x 100 m on the beds of Irish turloughs (temporary lakes that fill and drain through underlying limestone karst).
Survival of the algae, themselves, to prolonged exposure to drying is typically via modified vegetative cells with thickened walls, mucilage sheaths, and an accumulation of oils. The ability to resist exposure by such means is the major factor controlling zonation of algae at pond margins. Taxa with a gelatinous structure predominated in an intermittently dry section of Speirs' Pond in southern Ontario, and proved to be a subset (22) of those genera (30) found in permanently inundated sections (Williams et al. 2005). In the temporary site, green algae (Oocystis, followed by the filamentous Oedogonium) were dominant early in the hydroperiod, although diatoms (Pinnularia) were also present (Figure 4.3). For much of May, diatoms (Pinnularia, Navicula, and Synedra) predominated, including in the pre-summer- drought community—along with the Chlorophyta (Chlorococcum), Euglenophyta (Euglena), and some Xanthophyta (Tribonema). The community found in the mid-drought-respite period of late July (which resulted from heavy rainfall), consisted primarily of diatoms (Navicula, Diatoma), with some Tribonema (Xanthophyta).
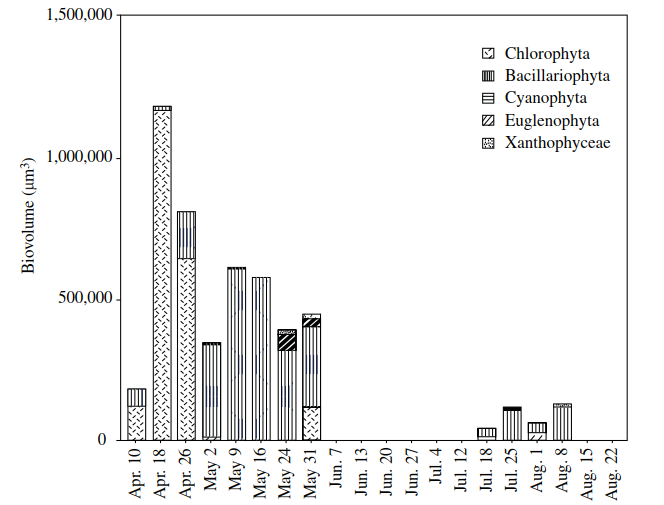
Figure 4.3. Temporal variation in the biovolume of major algal groups in an intermittently dry section of Speirs’ Pond, Ontario. The site was dry from June 7 to July 12, and again from August 15 to 22
Many species of alga in temporary ponds and streams appear to be opportunists. Many pass through predictable life cycle phases with maximum zygospore germination ocurring when water levels are highest. Vaucheria, a typical temporary pool alga, survives drought as the thick- walled zygotes discussed earlier, but it also has a 'back-up' system. The latter involves forming hypnospores in response to rapid desiccation. These are specialized structures that release amoeboid cells capable of movement to areas where water is more abundant and where they give rise to new filaments (Sands 1981). Such properties are clearly those of algal species adapted to temporary waters, as Benenati et al. (1998) found that the recovery and maintenance of the phytobenthos in a permanent, but regulated, river in Arizona were compromised by erratic flow management. Repeated desiccation of the algal community had major effects on the bottom-up interactions of the Colorado River ecosystem.
A number of algal species live epizootically on the branchiopod crustaceans found in temporary ponds. In Moroccan ponds, Thiery and Cazaubon (1992) found that filamentous greens, such as Stigeoclonium and Oedogonium, predominated on the shells of conchostracans (Spinicaudata), whereas small species of Chlorococcales and Tetrasporales colonized anostracan bodies. All of the algae were common species with wide hol- arctic distributions, such that no special adaptations were evident, and colonization site was thought to be determined by the microhabitat and swimming behaviour of the hosts.
Protozoans. Few studies have tackled the protozoan component of temporary water communities. Fenchel (1975) found densities of around 106 m~2 and biomass of 20-40 mg m~2 for ciliates in an arctic tundra pond between June and August. Some fed on algae or bacteria, whereas others were carnivores feeding on zooflagellates and other ciliates. Stout (1984) studied the protozoans of seasonally inundated soil under grassland in New Zealand and found a resemblance to the biota of a shallow temporary pond: an autotrophic community of phyto- flagellates and diatoms, and a largely bacter- iovorous community of zooflagellates, sarcodines (33 species), ciliates (57 species), and meiofauna. Of the 94 species of protozoan recorded, 45% could be described as freshwater forms and 35% as aquatic-terrestrial, the former dominating during winter when the soil was covered by several centimetres of water.
In an experimental ricefield, in Italy, Madoni (1996) found that the ciliate community was strongly influenced by environmental factors generated by the growing rice plants. Initially, when phytoplankton productivity was high both in the water column and at the sediment-water interface, grazing benthic species dominated the system. However, as the rice plants grew, algae decreased (likely due to shading and competition for nutrients) and decomposition processes became more intense. In the water column, this led to a predominance of bacteriovorous ciliate species, whereas the shift towards reducing conditions at the sediment-water interface resulted in the disappearance of many species and the relocation of others. In the final phase of rice cultivation, general ciliate diversity and production decreased markedly. Interestingly, despite these changes, one species, the prorodontid Coleps hirtus, dominated the system throughout the 4-year study—this was attributed to its extreme flexibility in feeding behaviour which allowed it to ingest algae, bacteria, other protozoans, and even small metazoans.
Coleps hirtus was also a dominant species in Laird's snow-melt pond in northern Quebec. Together with nine other common species, it formed the first phase of this pond's ciliate community. Using the 'Saprobiensystem' of classifying fresh- waters by their degree of organic enrichment, Laird found that the post snow-melt, oligosaprobic phase of the pond was quickly replaced by beta- mesosaprobic conditions which lasted about one week. Subsequent high bacterial production resulting from decaying grasses produced alpha- mesosaprobic conditions which, after another week, gave way to polysaprobic conditions. Ciliates found in these three enrichment phases of the pond are listed in Table 4.2.
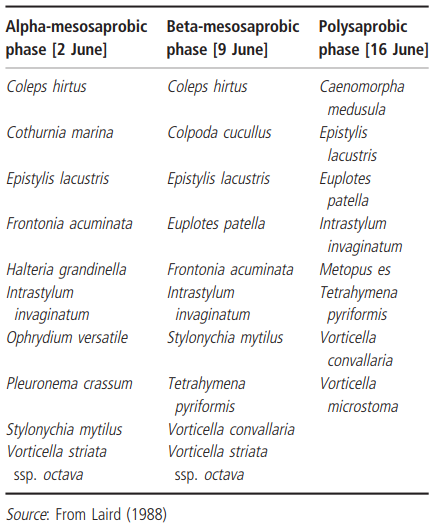
Table 4.2. Succession of the most common ciliate species throughout three enrichment phases in an intermittent, snow-melt pool in northern Quebec
Few studies have addressed inter-pond comparisons. However, Andrushchyshyn et al. (2003) showed that the relative abundance of ciliates in two adjacent intermittent ponds in southern Ontario differed significantly from one another: that in Pond II rose rapidly from day 1 increasing two orders of magnitude by day 7. In contrast, abundance in Pond I began at the same level but increased much more slowly, reached a plateau of around 500 individuals l_1, and increased again late in the hydroperiod (Figure 4.4(a)). The two ponds were also fairly dissimilar in terms of their species richness and species composition. Pond I contained 50 species compared with 70 species for Pond II, with only 24 species shared. Variation in ciliate abundance in Pond I could be explained by the number of days after filling (39%) and enclosure treatment (23%).
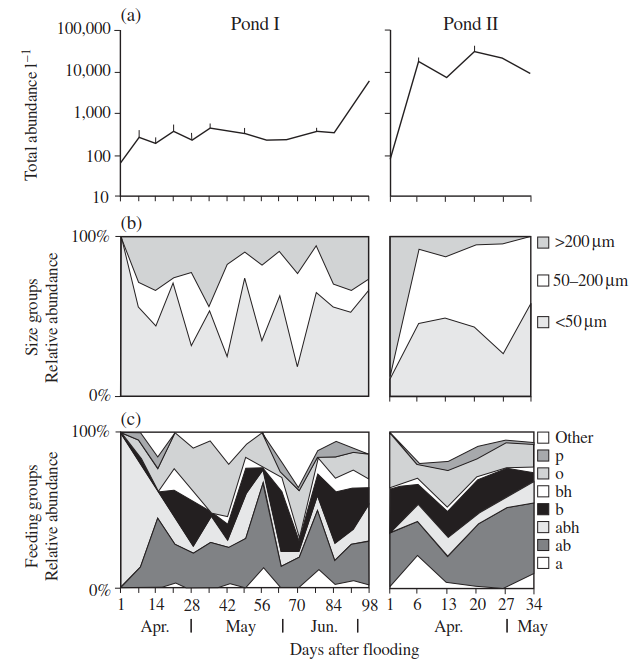
Figure 4.4. Seasonal development of (a) total ciliate abundance in two intermittent ponds in southern Ontario (means ±1 SE); (b) three size groups within the two communities; (c) relative abundance of the dominant ciliate feeding categories (a = algivores; b = bacteriovores; h = predators on heterotrophic flagellates; p ¼predators on other protozoans, including ciliates; o = omnivores; other = unknown prey)
These two parameters also explained 72% of the variation in species richness in Pond I. Sixty-five per cent of the variation in abundance in Pond II could be explained by the number of days after filling (27%), pH (19%), and nitrate levels (12%). Fifty-two per cent of the variation in species richness was explained by environmental parameters, of which pH was the most influential. Species succession was a strong feature of both ponds. Pond II contained more mid-sized ciliates (50-200 pm), whereas Pond I was dominated by smaller ciliates, especially in mid-May and early June (Figure 4.4(b)). There were more algivorous species in Pond II, although their abundance was greater in Pond I. In Pond II, bacteriovore relative abundance increased as the pond dried up; in Pond I there was a similar but more variable trend. Facultative algal feeders were most numerous in Pond II early in the season (e.g. 38% of the relative abundance on day 7 was due to Uroleptus gallina alone) but thereafter decreased rapidly. Pond I contained only one, though very abundant, facultative algivore, Pela- gohalteria cirrifera. Omnivorous ciliates were moderately important parts (21%) of the communities of both ponds early in the season but in Pond I thereafter declined (Figure 4.4(c)).
Although 13-14% of all species in the two ponds were predators, feeding chiefly on other ciliates, their abundance never exceeded 5% (Pond I) and 8% (Pond II) of the total ciliate community. Predator peaks occurred at around the same time (days 13-21) in both ponds. Experimental addition of invertebrate predators to these ponds resulted in higher ciliate abundance and species richness for a limited time in one of the ponds—suggesting that differences in food-web dynamics may influence ciliate community composition.
Stout (1984) put forward the idea that the biotas of moist soils and temporary ponds lie on a transition from freshwater to edaphic habitats— indeed, in a review of the world soil ciliate fauna, Foissner (1998) concluded that of the, at least, 1,000 known species about 25% are also known from freshwater habitats. Protozoan faunal elements from each can persist in the restricted moisture films around soil particles in pond bottoms, and extend into forest litter where physical contact between fallen, decaying leaves provides an unbroken moisture film. From here, Stout hypothesized that protozoans and some meiofauna have invaded soils, root zones, and vegetation surfaces, and have even been provided with colonization pathways to aerial habitats such as tree holes, leaf axils, and pitcher plant chambers (Figure 4.5).
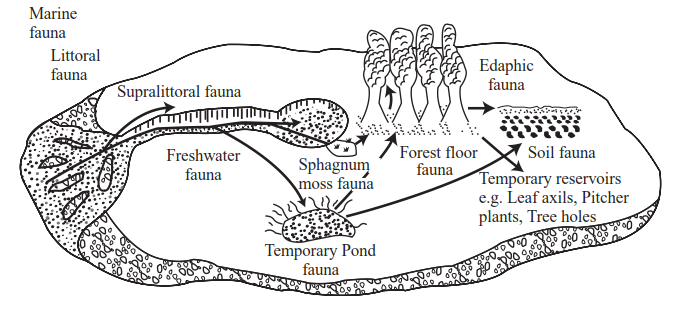
Figure 4.4. Seasonal development of (a) total ciliate abundance in two intermittent ponds in southern Ontario (means ±1 SE); (b) three size groups within the two communities; (c) relative abundance of the dominant ciliate feeding categories (a = algivores; b = bacteriovores; h = predators on heterotrophic flagellates; p = predators on other protozoans, including ciliates; o = omnivores; other = unknown prey)
Fukada et al.'s (1997) survey of Floridian container habitats revealed four protozoan parasites of mosquito larvae, the most common being Asco- gregarina taiwanensis (Apicomplexa) and the microsporidium Vavraia culicis.
Date added: 2026-07-14; views: 5;