Fungi and Plants in Temporary Waters: Ecology and Adaptations
Fungi. In running waters, aquatic hyphomycetes are known to be critical to the breakdown of riparian leaf litter, as well as serving as an important food source for many benthic invertebrates—indeed they are fundamental to the base of lotic food webs (Kaushik and Hynes 1971). It is likely that they are also important contributors to nutrient release and dynamics in temporary waters, especially, for example, those nutrients contained within the terrestrial summer vegetation that grows in dry pond basins and stream channels. Unfortunately, little work has been done on these habitats. However, in a study of an intermittent vernal pond in southern Ontario, Barlocher et al. (1978) found the fungal flora on bottom detritus to be very different from that found in nearby permanent ponds. Terrestrial fungi predominated during the waterless period and a seasonal succession was evident. In running waters, fungi actually add protein to decomposing leaves, a process that has since been described as 'conditioning' (Barlocher 1985).
Detritivorous insects, when given a choice, select conditioned leaves over those that are fresh, because of their enhanced nutritional value. The mechanisms used by fungi to degrade leaves have been examined by Suberkropp and Klug (1980), by initially isolating five species of aquatic hyphomycete (Alatospora acuminata, Clavariopsis aquatica, Flagellospora curvula, Lemonniera aquatica, and Tetracladium marchalia- num) and growing them in pure culture with hickory leaves. Enzymatic activity of each fungal species resulted in the skeletonization of leaves via maceration of the leaf matrix and the subsequent release of leaf cells as FPOM. After incubation, further fractionation and analysis of leaf material indicated that all fungal species metabolized (degraded) cellulose, and that two species (T. marchalianum and F. curvula) metabolized hemicelluloses. In cultures treated with T. marchalianum, the release of fine particulates coincided with increases in fungal biomass (as measured by ATP) and enzymatic activity in the supernatant, which degraded carboxymethycellulose, xylan, and polygalacturonic acid. Macerating activity increased with pH, indicating that pectin transeli- minase was involved in the softening of leaf tissue by T. marchalianum. This finding prompted Suberkropp and Klug to suggest that transeliminase is more important in the release of leaf cells than hydrolytic enzymes with lower pH optima.
Aquatic insects show a positive selection for conditioned food. Bacteria also can condition detrital material, but evidence suggests that fungi may be more effective in enhancing its nutritional value. For example, when Mackay and Kalff (1973) presented leaf discs from fungal and bacterial cultures to the caddisflies Pycnopsyche gentilis and P. luculenta, it was the fungal discs that were preferred.
Higher plants.As previously indicated, most temporary ponds represent the penultimate stage of a sere, the climax of which is terrestrial. In a study of plant succession in temporary ponds in Oregon, USA, Lippert and Jameson (1964) found that the species present were characteristically those found in wet places, for example, the cat-tail (Typha latifolia) and the spike rush (Eleocharis palustris). In northern Germany, Caspers and Heckman (1981) found T. latifolia and the grass Glycera maxima in large numbers in Stage 5 (the final stage) of their ditches. In this locality, these latter two species are eventually responsible for the total disappearance (terrestrialization) of the aquatic habitat. The margins of temporary waters are very susceptible to invasion by terrestrial species, the roots of which may proliferate through the basin substrate. Litter from all species provides a rich substrate for microorganisms and subsequently a source of food for aquatic invertebrates. Barlocher et al. (1978) measured an average of 132.8 gm~2 of leaf litter (ash-free dry weight) entering Ontario pond basins in the autumn.
Many temporary waters, especially those with longer hydroperiods and low- or no-flow, support heavy growths of aquatic macrophytes. In certain localities, free-floating annual macrophytes such as Azolla may contribute significantly to the productivity of temporary ponds. The benthic invertebrate communities of shallow lakes on the floodplain of the River Parana, Argentina, experience shifts in composition related to floating macrophyte cover and changes in depth. During high-water periods, communities are similar among lakes and are dominated by ostracods and oligochaetes. During low water, when biomass is highest, chironomid larvae (Chironomus) and burrowing mayflies (Campsurus) have been found to dominate lakes with 60% cover of Eichhornia cras- sipes, whereas lakes without floating plants were dominated by sphaeriid clams (Pisidium and Eupera), snails (Littoridina), and burrowing mayflies (Bechara 1996). Macrophytes provide an important structural 3-dimensional framework for the activities of many invertebrates, extending the benthic realm into the water column, and perhaps allowing greater interactions between bottom- dwellers and the plankton. However, some invertebrates also eat plant parts, especially the leaves. Cronin et al. (1998) found that aquatic insects consumed 0.2-1.7% of the leaf surface of water lilies (Nuphar variegata and Nymphaea odorata) per day in a Michigan lake, with some leaves suffering more than 60% damage. Submerged leaves appeared to be more susceptible. Interestingly, their data also provided support for the hypothesis that herbivorous insect species belonging to primarily aquatic groups have a much wider diet than those derived from terrestrial groups.
In a study of the microdistribution of macrophytes of vernal pools in southern California, Zedler (1981) related water level to a succession of species groups each with characteristic germination times, and flowering and seed maturation stages. Species were assigned to weighted average water duration classes (WADC), calculated from duration-frequency data, ranging from 1 to 16. Species falling into classes 1 to 4 are those rarely found within the inundated areas of pools but which grow commonly near the margins (e.g. Bromus spp., bromegrass; and Erodium spp., filaree). Species with WADC values of between 4.5 and 7.0 (e.g. Juncus bufonius, toad rush; and Agrostis microphylla, bentgrass) possess a physiology capable of tolerating inundation but fall short of being true marsh or aquatic plants. Species in classes 7.0-9.5 tolerate water-cover for long periods, but do not thrive under long submergence— they are able to germinate and grow underwater but require a considerable period out of water in order to mature (e.g. Callitriche marginata, water- starwort; and Anagallis minimus, false pimpernel). Characteristic temporary pool species fall into the class range 9.5-11.5. These are species possessing the morphological and physiological plasticity that allows them to withstand prolonged submergence (e.g. Downingia cuspidate; Pogogyne abramsii, pogo- gyne; and Eryngium aristulatum, marsh eryngo). All three species produce submerged leaves that are very different from the emergent foliage of the mature plant. The final group of plants (classes 11.5-16) contains species that are almost true aquatics, yet they can withstand some dry period (e.g. Pilularia americana, pillwort; Callitriche longipedunculata, water-starwort; and Lilaea scilloides, flowering quillwort).
On the basis of this study, Zedler proposed several hypotheses: The first was that the distribution of standing water in time and space is the single most important factor influencing temporary pool macrophytes. Physical stress of inundation is the primary cause of the distinctive assemblages of species but pattern of soil moisture may be important also. That waterlevel fluctuation stress differentially affects semi-emergent plants, can be illustrated by the water buttercup, Ranunculus. In a comparison of four species living on floodplains, He et al. (1999) showed that each exhibited different adaptations to inundation: R. repens, a species from lower, frequently inundated floodplains, proved to be very tolerant of prolonged waterlogging and submergence, as it was able to use oxygen generated by underwater photosynthesis for root respiration; R. sceleratus, common in low-lying mudflats, responded by ameliorating flooding stress through high root porosity and an ability to rapidly elongate its petioles under water; R. acris, from less- frequently flooded areas, responded in a manner similar to R. repens, although it had a lower resistance to submergence in the dark; and R. bulbosus, a species from seldom-flooded river levees, was generally intolerant to both waterlogged soil and complete submergence. Coxon (1987) similarly found that flooding depth was pivotal in the development of vegetation in Irish turloughs. Floor vegetation became increasingly dominated by marsh plants as the depth increased to around 3 m. Surprisingly, some of the deepest turloughs, along with those with variable depth due to undulating beds, had short hydroperiods, and a floor vegetation dominated by dry land ruderals (e.g. Rumex). Vegetation was, in addition, related to deposit characteristics, although the latter were also correlated with hydroperiod—peat and marl were associated with longer water residence, whereas sand, silt, clay, and diamicton were more common in turloughs with shorter hydroperiods. Linhart and Grant (1996) pointed out that soils are especially strong agents of selection in short-lived plant species. Consequently, pools located in regions with different soil characteristics (e.g. mineral content, pH) would be likely to harbour genetically differentiated populations of a variety of species.
Zedler's second hypothesis was that local extinctions of temporary pond species are rare despite considerable year-to-year variations in rainfall and habitat availability. Several characteristics of temporary pool species lessen the probability of extinction, namely small minimum plant size; small seed size and thus many seeds/unit biomass; and high vegetative and reproductive plasticity typical of annuals. Holland and Jain (1981), however, in a survey of over 250 vernal pools in central California, showed that species composition varied significantly among years and that species richness varied nearly twofold over sites, apparently in response to regional differences in rainfall among sites and edaphic conditions, and was less in a drought year. However, Gafny and Gaisith (1999) demonstrated that even though the historical appearance of macrophytes in the littoral zone of Lake Kinneret, Israel was sporadic (due to water level fluctuations), species stands were always located in exactly the same places, and that this was highly dependent on sediment structure. Husband and Barrett (1998) found that in the arid region of northeastern Brazil, the annual Eichhornia paniculata occurs in discrete temporary waters, such as pools, wet ditches, and flooded pastureland. Population size averaged 86 plants, with 64% of the populations persisting from one year to the next, and, on average, 21.6% of suitable habitat patches were occupied. No populations occurred in areas where the density of patches was less than around 2 patches km-1, suggesting the existence of a habitat threshold for persistence.
The third hypothesis was that despite the fundamental influence of inundation on community structure, competition is also a factor. Most temporary pool species are restricted at the upper (drier) end of the water duration-elevation gradient by competition from species that are better at exploiting drier conditions. At the lower (wetter) end, however, morphological and physiological tolerance to inundation are probably more influential. Holland and Jain showed that several grassland species grew in vernal pools during a year of severe drought but they were excluded in years of average rainfall. Further, several taxa characteristic of the vernal pool flora in this region were not evident during the drought although they were common and widespread in subsequent years. They showed that the number of species specialized for a given pool depth is proportional to the relative area in a pool at that depth; thus more species are adapted to the margins than to the centre. They concluded that species richness is determined chiefly by certain physiographic niche properties whereas competitive niche-partitioning factors influence congeneric sympatry and zonation within a pool.
Elam (1998) has reviewed the limited population genetics data of vernal pool plants and concluded that species often form 'races' or biotypes, at the geographical (among-population) scale, but also show intrapopulation differentiation at the local scale. The latter may involve changes over distances of as little as 2-5 m. In Veronica peregrina, for example, the environmental conditions experienced by individual plants from a pool's centre to its margin are known to change with respect to competition regime, moisture availability, and environmental predictability (Linhart 1988). At the centre, individual plants grow among a high density of conspecifics, but under a relatively predictable, wet environment. In contrast, individuals growing at the margin come under heavy interspecific competition from grassland species, as well as having to endure a low moisture, less predictable environment. The resulting differentiation occurs over distances not considered a barrier to gene flow by seed and pollen, and suggests that the homogenizing process of gene flow can be offset by the effects of strong disruptive selection (which favours extreme traits in a population when sudden changes occur in the environment).
Local climate change can influence the macrophytes associated with shallow ponds. For example, the mid-1990s in Britain were considered to be drought years in which the vegetation of a pond in Epping Forest was invaded by creeping bent grass (Agrostis stolonifera), with isolated clumps of Yorkshire fog (Holcus lanatus) becoming established on the margins. In prior, wetter years (1989 and 1991), the dominant vegetation was floating sweet-grass (Glyceria fluitans), together with soft rush (Juncus effusus), reed mace (Typha latifolia), and yellow flag (Iris pseudocorus). Panter and May (1997) concluded that should the drying trend persist, the pond would be likely to continue to develop a terrestrial succession of plants. Welling et al. (1988) working on prairie wetlands showed, similarly, that if drought conditions were extreme this favoured the germination of annual species over emergents. Reflooding, on the other hand, eliminated annuals and stimulated the vegetative growth of emergent species, unless the latter's water depth tolerance was exceeded (Poiani and Johnson 1991). In the coastal wetlands of southeastern Alaska, Pollock et al. (1998) showed that sites with intermediate flood frequencies and high spatial variation of flood frequencies (SVFFs) were plant species-rich, whereas sites frequently, rarely, or permanently flooded and with low SVFF were species-poor (Figure 4.6). Such changes in plant species composition are likely to affect faunal abundance and composition. For example, Wrubleski (1987) demonstrated that the microdistributions of insects (Chironomidae) in Delta Marsh, Canada varied among major vegetation communities along a gradient from deeper water to dry land (Figure 4.7).
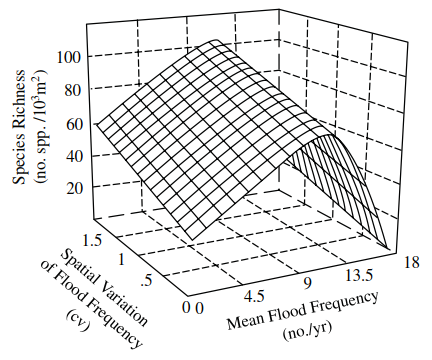
Figure 4.6. Three-dimensional representation of the relationships among macrophyte species richness, the spatial variation in flood frequencies, and mean flood frequency for 16 wetland sites on Chichagof Island, southeastern Alaska (redrawn after Pollock et al. 1998)
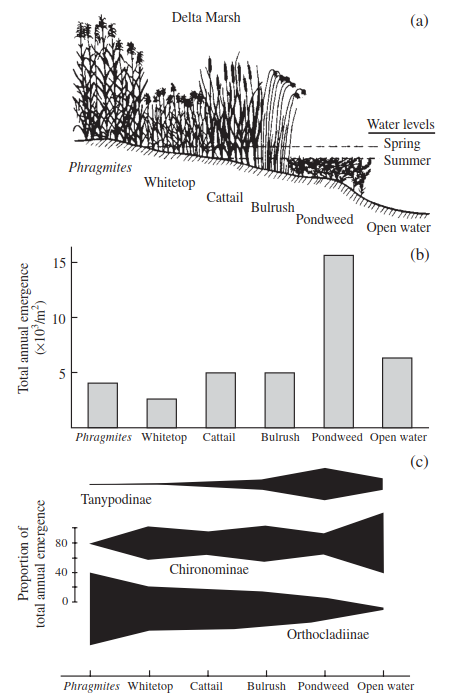
Figure 4.7. Microdistribution of chironomids along a moisture and vegetation gradient in Delta Marsh, Canada: (a) elevational and vegetation gradients; (b) total annual chironomid emergence from each microhabitat; (c) proportion of the total emergence represented by each chironomid subfamily (redrawn after Wrubleski 1987)
The Pantanal region of Brazil supports the largest wetland complex in the world, part of which includes vegetation well adapted to seasonal cycles of flooding and drought (Por 1995). Several vegetation zones have been identified, and those most associated with the network of water channels include: (1) Amphibious herbaceous vegetation that grows under permanently wet, temporarily submerged conditions, such as are found associated with shallow lakes (baias), and includes grasses (Cyperus giganteus, Scirpus validus, and Paspalum repens), the reed Typha dominguensis, and the maranthaceans Thalia geniculata and Polygonum hispidum; (2) Herbaceous campo vegetation that represents seasonally flooded grasslands, comprising species of Panicum, Paratheria, Oryza, and Setaria, together with some exotic herbs, such as Brachiaria, introduced by cattle ranchers; (3) Gallery forests that cover the higher banks of the major rivers and comprise Triplaris formicosa, Rheedia brasiliensis, several species of Inga, Vochysia, and Ficus, including Ficus elliotiana a species endemic to the Pantanal and especially well adapted to surviving drought; and (4) Cerrado-type forest (shrub savannah) that may often also cover seasonally flooded forest floors. However, whereas the reliance of these plant communities on inundation cycles is quite well understood, comparatively little is known of their influences, energetic or otherwise, on the aquatic communities.
In a review of the adaptations of plants to flooding stress, Blom (1999) emphasized that, for woodlands, zonation is largely determined by local hydrology. For example, successful establishment of softwood species, such as those belonging to the Salicaceae, on river banks depends on the interaction between water levels and the timing of seed dispersal—they are well adapted to irregular, high, and prolonged inundation. In contrast, hardwoods, such as those belonging to the genera Quercus, Fraxinus, Ulmus, and Acer, are flood- sensitive, are found higher on the floodplain, and have heavy seeds that germinate in shade. The most shade-tolerant hardwoods are the least well adapted to inundation. These properties will influence which species are likely to have their roots in and around temporary water bodies, and hence the quantity and quality of allochthonous leaf litter that will fall into the basin and upon which the aquatic food web will, in part, be based. Further, inundation-tolerant species survive as a result of having aerenchymatous roots together with shoot parts that elongate upon submergence. Most importantly, as a result of high porosity in the roots of inundation-tolerant species, radial oxygen loss greatly influences nitrification and denitrification processes in the flooded basin soil. Root-derived oxygen thus has the capacity to restore nutrient cycles and 'detoxify' the oxygenated rhizosphere.
Forestry practices, especially the stage in forest cycles, have been shown to influence low flow conditions in streams. For example, pre-planting drainage may increase low flow if more than 25% of the watershed is drained. In contrast, forest growth decreases low flow levels (in all but the driest years). Clearfelling increases low flows initially, but this may be followed by a gradual decrease depending on the rate of vegetation regrowth (Johnson 1998). As streams dry up, the degree of retreat of the groundwater table can have a profound effect on the structure of riparian plant communites. For example, in the San Pedro River floodplain, Arizona, Stromberg et al. (1996) observed that as the depth to groundwater ranged from 0 to 4 m, their wetland indicator score (an index based on cover of plants within wetland indicator groups and frequency of indicator species) changed sharply. In particular, the abundance of obligate wetland herbs declined significantly at groundwater depths below 25 cm. Other impacts of retreating groundwater included reduced establishment of Populus fremontii-Salix gooddingii forests, and reduced cover of herbaceous species associated with fine-textured soils and the shady conditions of floodplain terraces.
Forestry practices may also influence the structure of temporary water communities through alteration of shading. For example, woodland ponds are heavily shaded, have little emergent vegetation, and receive much of their energy input from fallen tree leaves—the communities that have evolved under these conditions are thus largely detritus-based. Timber harvesting removes this cover, opens the ponds to sunlight, reduces allochthonous energy input, and promotes a shift to alga/herbivore-based communities.
The presence of temporary waters in arid regions can have a profound influence on the local riparian flora. For example, in the western United States, where riparian areas occupy less than 1% of the landscape, Zimmerman et al. (1999) found that variation in vegetation distribution and composition in Arizona was best explained by a complex temperature/moisture/substrate gradient, with understory diversity being related to changes in slope and sand/gravel substrate. Most of these environmental properties are of course running- water-induced. Further, each tributary canyon supported unique and oftentimes rare plant species. In southern California, Bauder et al. (1998) identified five vernal pool plant species as being endangered, a consequence of a historical 95-97% loss in pool habitat.
Date added: 2026-07-14; views: 5;