Ecosystem Complexity and Interactions of Processes and Drivers
Ecosystems are, by definition, a complex network of relationships and processes—that is, they are determined by a multitude of factors and interactions. The complexity of an ecosystem develops very slowly over time, starting from a few actors—often mosses and lichens, such as on bare rock in glacier fore-fields when the ice retreats or after major disturbances such as eruptions of volcanoes—and eventually reaching the highly diverse ecosystems, such as temperate grasslands or tropical forests, that we know and use.
Thus, the age of ecosystems is difficult to determine, since many generations of plants, animals and microorganisms have shaped the ecosystems that we study. Furthermore, ecosystem development over time is not linear, but rates can change at varying stages over time, triggered both internally and externally. Sudden events or disturbances, occurring in minutes and hours, change ecosystems as fundamentally as slow continuous changes in environmental conditions, occurring over decades. Internal drivers of ecosystem change can be species competition and thus vegetation succession, but also accumulation and/or consumption of resources, which change basic life conditions. External drivers can be natural disturbances such as landslides or insect outbreaks, or anthropogenic impacts such as land use, climate change or introduction of invasive species. Moreover, the components of an ecosystem are able to adapt, self-organise and interact with each.
Unpredicted Existence of Neighbours. One of the most important emergent properties, from the point of view of an individual plant, is the unpredictable existence of neighbours, which can possess a multitude of different competitive traits. Thus, the success or failure of an individual plant or a species in an ecosystem is no longer solely determined by the physiological characteristics (as described in Part I) and responses of the species to the environment (as described in Part II), but is suddenly co-determined by random effects such as the presence or absence of competitive neighbours. Particularly at the early stage of stand development, the composition of vegetation is determined by germination rates, establishment and survival during the seedling stage. In the medium term, interactions among plant species become decisive.
In the longer term, mainly abiotic disturbance processes, sometimes also called interferences, become more important; plant responses to processes such as invasion, extreme events or management practices may drastically change ecosystems within a very short time, sometimes as short as minutes or hours (e.g. hailstorm, flooding). Element or substance pools that have accumulated over centuries and millennia may disappear or degrade within a very short period, in turn also affecting the presence or absence of certain neighbours.
Scaling up of Processes. Processes within ecosystems occur at different spatio-temporal levels (sometimes also named scales), i.e. from foliage to landscapes or from seconds to millennia from small to large scales, respectively, also differing among components of the ecosystem. Often processes differ profoundly on different scales. For example, while small and fast scales are often dominated by chemical and biophysical processes, larger and slower scales are rather controlled by interspecies competition, species composition and community ecology. On very large scales, environmental boundary conditions affect ecosystem structures and dynamics over centuries and millennia. Nevertheless, even if these processes are identical (for example, photosynthetic CO2 fixation) the response of the process to the same driver (in this case, light intensity) will differ with the organisational level or scale—that is, from leaves to fully leaved branches to the entire ecosystem.
Thus, processes cannot simply be scaled from the leaf to the ecosystem level because additional constraints, such as spatial structure—an emergent property of an ecosystem—come into play. One classical example of such an upscaling issue is net assimilation. While leaf photosynthesis follows a saturation function driven by the availability of light and CO2, as well as vapour pressure deficit, photosynthesis at the ecosystem level is rather linearly related to incoming light. Moreover, it reaches much higher assimilation rates, in comparison with the leaf or branch levels, at higher light intensities—not reaching saturation— because more light can be used by more leaves within the entire ecosystem.
Moreover, the light compensation point is higher at the ecosystem level than at the leaf or branch levels because more light is needed to overcome auto- and heterotrophic respiratory losses of CO2 in low-light conditions within the entire ecosystem, in comparison with the leaf or branch levels (Fig. 13.1). The leaf area index (LAI) and thus light attenuation (i.e. the decrease of incoming light supply deeper in the canopy), as well as the nutrient supply, determine the actual magnitude of net assimilation at the ecosystem level.
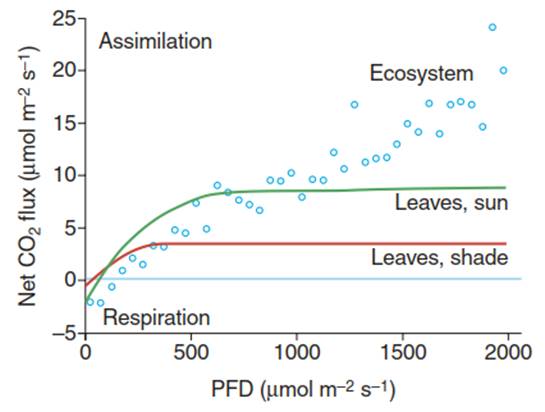
Fig. 13.1. Upscaling of CO2 exchange from leaves to the ecosystem. Differences in net CO2 fluxes—that is, assimilation and respiration—in response to light intensity (PFD, photon flux density) between foliage (beech leaves) and the entire ecosystem. (Leaves: after Larcher (2003); ecosystem: data from E. Paul-Limoges)
Thus, the emergent property of an ecosystem (here, ecosystem structure) can change the response to the same driver (here, light intensity) of an ecosystem in comparison with its components. These new characteristics can be related to resource use at the ecosystem level but can also be linked to the interactions of different species (Part IV).
Date added: 2025-02-05; views: 559;