Microorganisms and Trophic Structure
What role do microorganisms specifically play in energy flow and biogeo chemical cycling? They have not been conspicuous in typical food web diagrams and were largely overlooked until recently. In part this is because they are invisible and appropriate methods for assessing standing stocks and turnover in nature have been difficult to devise (DeLong and Karl 2005; Zinger et al. 2012). Actually, despite the focus on macroorganism-based grazer chains, most—and in some cases almost all—of the net primary production (NPP) in most natural ecosystems is utilized in the detritus webs.
Microbes are important in both grazer and detritus chains (Steffan et al. 2015), but especially in the latter, where they play the seminal role as decomposers of plant and animal carcasses, then are in turn eaten. They grow as parasites or commensals on the surface of or within plants and animals (Andrews and Harris 2000; Dethlefsen et al. 2006; Dobson et al. 2006; Lafferty et al. 2006) in both terrestrial and aquatic ecosystems. Many people associate aquatic primary production with macrophytes (submerged or emergent plants), yet most of it is generated by phytoplankton. For instance, a genus of photosynthetic cyanobacterium, Prochlorococcus, is the most abundant (global estimates ~1027 cells) photosynthetic organism not only in the oceans but on Earth and may account for up to 50% of NPP (Biller et al. 2015; for planktonic eukaryotes, see de Vargas et al. 2015). It is also the smallest free-living photosynthetic cell and contains the smallest genome. In his famous book on plant biology, Corner (1964, p. 3) has described the vast communities of oceanic phytoplankton imaginatively as .. the sea meadows of which the oceanographer now writes consist not of blades of grass but of separate cells, corresponding with those that build the blade of grass. They are not disintegrated plants, but plants too primitive for integration”.
Beginning in the 1970s, a paradigm shift occurred with respect to energy flow and nutrient cycles in the ocean. Initially, this was the recognition of the importance of a 'heterotrophic microbial loop' (Fig. 3.2) based on bacterial consumption of dissolved organic matter entering the water column from primary producers (Pomeroy 1974; Azam and Malfatti 2007; DeLong and Karl 2005; Karl 2007). A large portion of the organic matter synthesized by phytoplankton, photosynthetic bacteria, and seaweeds is exuded; heterotrophic microorganisms convert the dissolved organics to biomass that re-enters the conventional nanoflagellate → ciliate → mesoplankton → herbivore/carnivore levels (shorter routes are possible; Pomeroy et al. 2007). Bacteria also play a major role themselves in releasing organics, much of which is recalcitrant to further breakdown; these products are sequestered as particulate fixed C and sink into deeper waters.
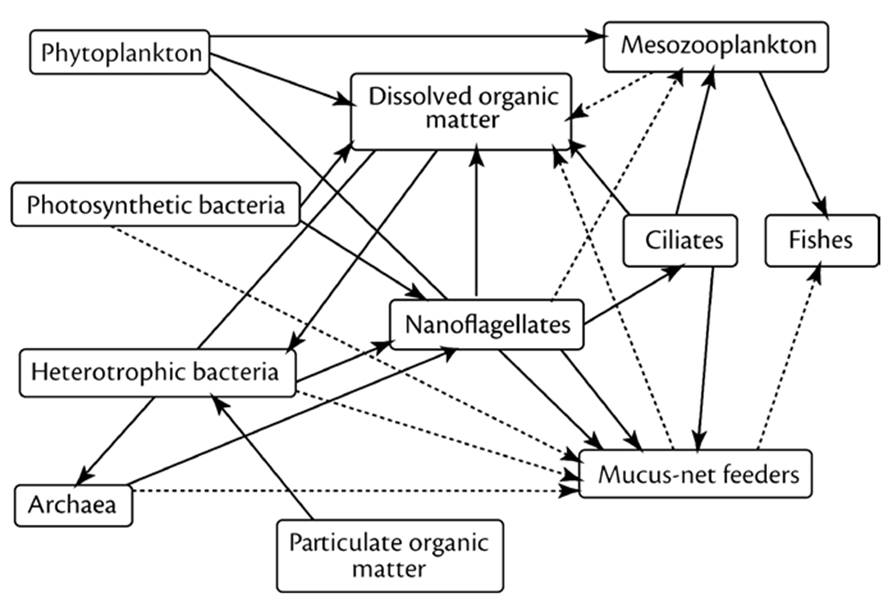
Fig. 3.2. Oceanic food web emphasizing the so-called “microbial loop”. All organisms shown except the fishes and mesozooplankton (top right side and including mucus-net feeders, bottom) are members of the loop. The continuous lines show major fluxes of energy and carbon; dotted lines show those of lesser magnitude. Note the central role of dissolved organic matter (box, top center of figure), which is utilized by Bacteria and Archaea, with the flux then moving sequentially through nanoflagellates, ciliates, mucus-net feeders or mesoplankton, and ultimately fish. A significant and in some cases the dominant portion of primary production is now interpreted to occur by mixotrophic protists (nanoflagellates and ciliates; see text and Mitra et al. 2014). From Pomeroy et al. (2007). Reproduced from Oceanography by permission of The Oceanographic Society © 2007
These discoveries are remarkable because the dogma for oceanic food webs had featured a strictly compartmentalized grazer chain for C flow based on consumption of phytoplankton by ciliated protozoa followed by copepods, which were in turn eaten by progressively larger consumers. Now it is recognized that about half or somewhat more of the primary production of oceans passes via the microbial loop, which has a broad influence on marine biogeochemical cycles (Azam and Malfatti 2007). In oceans, the dominant life forms in terms of numbers, species diversity, genetic diversity, and metabolic activity are bacteria.
The second and equally exciting part of the paradigm shift was recognition of a major segment of the plankton community that operates as mixotrophs. Mixotrophy refers to the combined use of light energy to fix C autotrophically along with heterotrophic acquisition of organic carbon sources (and other organic as well as inorganic nutrients). Previously, mixotrophy was relegated to a few biological curiosities such as carnivorous plants that photo - synthesize but also capture and digest insects. It became apparent that many protist species (dinoflagellates and ciliates) in marine ecosystems either have evolved this ability constitutively, or acquire photo trophy in various ways (Flynn and Hansen 2013). Frequently, this is phagotrophic when they capture algal prey, which eventually become endosymbionts, or by digesting the prey and retaining their chloroplasts (Stoecker et al. 2009). Mixotrophy not only changes fundamentally the structure of ocean biogeo chemical cycles (Mitra et al. 2014) but means functionally that eukaryotic plankton across many size ranges can survive competitively in nutrient-deprived (N, P, Fe; Moore et al. 2013) surface waters. This flexible nutrition strategy appears to enable mixotrophs to dominate the plankton community under certain conditions by outcompeting strict phototrophs (Hartmann et al. 2012; Ward and Follows 2016).
Another component of this ongoing trophic reinterpretation occurred in 2000, prompted by a genomic analysis of the bacterial component of the plankton community. Among the marine bacterioplankton are forms that can obtain their energy as nonphotosynthetic phototrophs (Beja et al. 2000; see also Venter et al. 2004). (Strictly speaking, these microbes are not photosynthetic because they lack chlorophyll and do not engage in the classic photosynthetic reaction typical of photolithoautotrophs.) They capture solar energy by utilizing a membrane protein called proteorhodopsin, a form of the visual pigment rhodopsin that functions as a light-driven proton pump and coupled photophosphorylation. Interestingly, these bacteria also grow as chemotrophs as has been demonstrated by growing them in culture in darkness, where they use organic carbon as an energy source. One genus of bacterium that lives this way is Pelagibacter, but ongoing analysis of pelagic (water column in the open ocean) waters by environmental genomics and direct gene probes indicates that as many as half of the upper ocean’s bacteria may harbor this photoprotein. Laboratory experiments under light/dark and high/low nutrient conditions with marine Vibrio suggest that they have a fitness advantage over competitors in the frequently occurring low nutrient (oligotrophic; see terminology below) oceanic environment by supplementing conventional heterotrophy with light energy (DeLong and Beja 2010; Gomez-Consarnau et al. 2010). To date none have been found that fix carbon, so technically they are not autotrophs but rather photoorganoheterotrophs.
A final climax in this remarkable series of discoveries is that work beginning in the 1970s documented novel deep-sea ecosystems (reviewed by Van Dover 2000; Kelley et al. 2002; Cavanaugh et al. 2013). These diverse marine invertebrate communities, both in relatively cold water and surrounding deep-sea thermal vents, are based on the productivity of lithotrophic bacteria, which oxidize H2S or other inorganic substrates such as H2 and CH4. These substrates, acting as electron donors (as well as the electron acceptor CO2), originate in the hydrothermal fluid. Thus, we now know that there are not one but two major systems for primary production on Earth: While solar radiation provides energy for phototrophy, terrestrial thermal energy and pressure provide the energy source on venting water for anaerobic or aerobic chemosynthesis by the lithotrophic reduction of CO2.
To put the chemosynthetic system in perspective, however, it should be noted that current estimates indicate (Van Dover 2000; McClain et al. 2012) that sinking particulate matter representing carbon originally fixed by photosynthesis in the upper reaches of the ocean contributes by far the greatest percentage of chemical energy for deep-inhabitants; chemosynthesis contributes only about 0.02-0.03% total ocean production and about 3% of the carbon flux to nonchem- osynthetic organisms. These numbers may well be revised upward as further such communities become discovered and explored.
Date added: 2025-06-15; views: 351;