Complex Life Cycles. Concept and Definition
Concept and Definition.The terminology and theory of complex life cycles (CLCs) were formalized largely by zoologists to describe life cycles in which there is “an abrupt ontogenetic change in an individual’s morphology, physiology, and behavior, usually associated with a change in habitat” (Wilbur 1980; p. 67). An organism that passes through two or more such (irreversible) distinct phases was considered to have a complex life cycle (Istock 1967). In contrast to these complicated cycles, if the offspring of a species are born into essentially the same habitat occupied by the adults, and do not undergo sudden morphological or ontogenetic change, the organism is said to have a simple life cycle (SLC). Examples of the latter include birds, humans, and other mammals.
Animals The classic example of a CLC is the frog, many species of which occupy two distinct ecological niches: that of an aquatic herbivore in the tadpole phase, followed by life as a terrestrial carnivore in the adult stage. As Wilbur notes, it is interesting that two such species may be ecologically and morphologically more similar as tadpoles than either is to the adult frog it will become. CLCs are represented in most animal phyla, have evolved independently many times, and have persisted through geological time (Moran 1994). Examples include in general the amphibians, insects (especially aquatic insects, which live for months or years as immature forms in streams, followed by an ephemeral adult phase of hours to weeks on land), and most marine invertebrate species such as barnacles, starfish, snails, etc., that have a tiny larval phase that feeds in coastal waters followed by an adult phase on rocks or in mud (Fig. 6.4a and Roughgarden et al. 1988). Where the individual organism passes sequentially from larva to adult by metamorphosis the type of CLC is single-generational (as opposed to multigenerational, below).
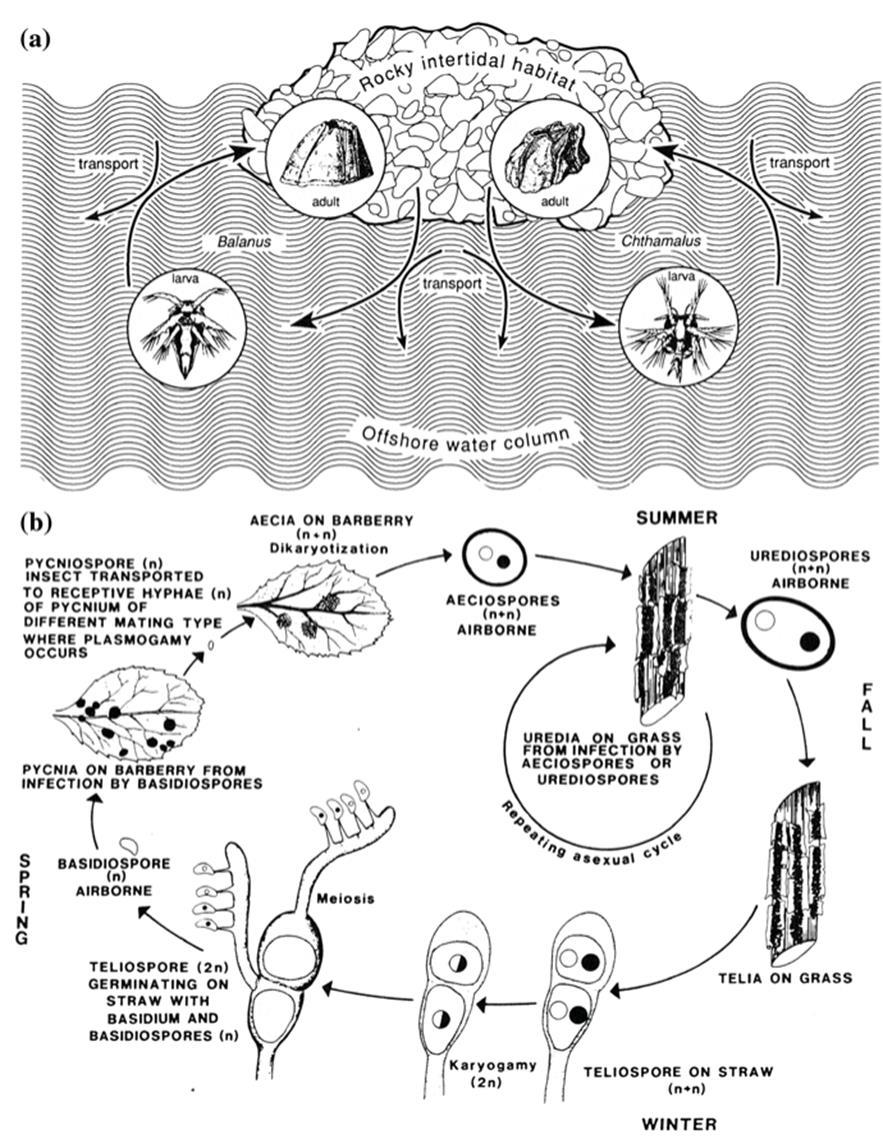
Fig. 6.4. The complex life cycle as it pertains to macroorganisms and microorganisms. a The tiny, motile larval phase of barnacles (e.g., Balanus and Chthamalus) lives in offshore waters; the sessile adult phase occupies the rocky intertidal zone. Figure redrawn, based on Roughgarden et al. (1988); barnacles reproduced from Darwin (1859). b The rust fungus Pucciniagraminis alternates between the stems of grasses (cereals) and the leaves of barberry, with multiplication phases on each.
The five spore stages clockwise from the bottom right-hand corner are the teliospore, basidiospore, pycniospore (spermatium), aeciospore, and urediospore (urediniospore) with associated nuclear stages shown as n, n + n (dikaryotic), and 2n. For contemporary terminology and life cycle details, see Webster and Weber (2007, Chap. 22). From Roelfs (1985). Reproduced from The Cereal Rusts. Vol. II. Diseases, Distribution, Epidemiology and Control, A.P. Roelfs and W.R. Bushnell (Eds.), by permission of Alan Roelfs (Univ. Minnesota) and Academic Press, ©1985
Algae While zoologists played a seminal role in developing the conceptual framework of CLCs, clearly organisms other than animals exhibit an analogous pattern of drastic morphological and ecological change during the course of their life histories. The seaweeds and bryophyte land plants typically have a pronounced alternation of generations consisting of a prominent gametophytic phase contrasting with morphologically distinct sporophytes, discussed earlier. The heteromorphic algae, of which the red, brown, and green seaweeds are good examples, have two separate, ecologically distinct phases that may be so dissimilar morphologically that the phases have even been classified as separate species. For instance, in the red alga Porphyra, a haploid thallus of leafy sheets about 3-15 cm long growing on rocks in the intertidal zone alternates with an inconspicuous, shell-inhabiting sporophyte, the Conchocelis phase.
Frequently, the morphological and ecological divergence within the algal life cycle parallels the alternation of haploid with diploid generations. In some cases, the phases are self-perpetuating and neither morphological nor genetic alternation is obligatory (Lubchenco and Cubit 1980). Even in cases where the phases are morphologically similar, ecological differences likely occur and may explain in part the origin and maintenance of biphasic life cycles (Hughes and Otto 1999) noted previously. Instances of such isomorphy technically do not rise to the level of a strictly defined CLC. The heteromorphic algae, certain insects such as aphids that display different phenotypes during a seasonal cycle, many parasites, and the fungi discussed below, are examples of multiple generation type CLCs.
Fungi A situation analogous to the bryophytes and heteromorphic algae is the filamentous fungi. Members of the largest group of fungi (at least 30,000 species in the phylum Ascomycota) have diverse lifestyles but typically have two distinct, alternating forms, a perfect or teleomorph (sexual) state and an imperfect or anamorph (asexual) state. Each has its own characteristic structures, spore type and frequently, habitat (Fig. 6.4b). Many ascomycete species are associated with living plants both above and below ground as parasites, mutualists, or commensals. In terms of their ecological activities, these and other filamentous fungi could be conceptualized as passing through two life phases in which biomass is allocated primarily to either foraging or reproduction.
The former, a juvenile vegetative condition, is characterized usually by a diffuse hyphal network adapted for exploration and nutrient acquisition upon or within a substratum. The latter, a mature reproductive condition, involves initially specialized hyphae and ultimately propagules produced asexually, usually through repetitive cycles and released into the medium (air, water, or soil). Upon death of the plant or plant organ, the fungus frequently continues to grow as a saprophyte, undergoes sexual reproduction, and enters a relatively dormant condition until meiospores are released upon the appropriate environmental cues. In summary, although there is no host alternation here (unlike the rust case below), with this basic type of cycle there are morphologically and functionally distinct phases and two distinct niches. This pattern is consistent with Wilbur’s and Istock’s criteria for organisms with a CLC.
Animal and plant parasites. Among the most complex life cycles are those of many animal and plant parasites. These include the rust fungi reviewed at length below and many of the nematodes, trematodes, protozoa, and phytophagous insects that must use two or more hosts in sequence (Chaps. 2 and 3 in Price 1980; Thompson 1994). Usually the sexual phase of the parasite life cycle is associated with a particular host or habitat while the asexual stage, functioning mainly to increase and disperse offspring, occurs with a different host. The case for parasites can also be equivocal as one must distinguish complexity in the epidemiological sense from that of the life cycle of the causal organism. Many parasites attack a single host species. Although the epidemiology of the resulting disease may be complex, it does not follow that the incitant has a CLC.
In other cases, several species may potentially be hosts, yet neither is their sequential involvement mandatory for completion of the parasite’s life cycle nor does the parasite have to undergo an irreversible ontogenetic change. The epidemiology of scrub typhus presents a fascinatingly intricate case that illustrates the above points.
The concurrence of events culminating in human infection depends on: (i) presence of the bacterial pathogen Rickettsia tsutsugamushi; (ii) the vector, chiggers (trombiculid mites); (iii) small mammals, particularly rats, on which the chiggers feed; and (iv) transitional or secondary forms of vegetation (‘scrub’), through which humans chance to pass. The intimate relationship of these four factors led to the descriptor ‘zoonotic tetrad’ (Traub and Wisseman 1974). However, chiggers are the primary reservoir of the causal organism and the rickettsia need not pass through either rodents or humans to complete their life cycle. This rickettsial cycle is not complex in the ecological sense of Istock and Wilbur.
In contrast to typhus, the malarial parasite Plasmodium must pass through both the mosquito and human to complete its CLC, during which it undergoes alternation of generations and several morphological states. Similarly, the liver fluke (class Trematoda) has a CLC by virtue of obligately passing through snails and humans (or usually another vertebrate such as dogs, cats, pigs, or sheep) to complete its life cycle. In the process, the parasite passes sequentially from zygotes to ciliated larvae (miracidia), sporocysts, nonciliated larvae (rediae), tadpole-like larvae (cercariae), metacercariae, young and finally adult stages from which are produced the zygotes.
Date added: 2025-06-15; views: 523;