History of Vegetation to the End of the Tertiary
In the Precambrian Period two large separate landmasses existed, each near the poles: the Laurasia continent in the Northern Hemisphere, and the Gondwana continent in the Southern Hemisphere. These two continents over time merged and became the continent of Pangaea.
Pangea remained one mass until the Palaeozoic Period, where changes in positions towards the poles and equator occurred. Traces of life and the first single-celled prokaryotes (bacteria, cyanobacteria) existed for about three billion years in an oxygen-poor atmosphere. Eukaryotes and multicellular forms could only develop when increases in oxygen occurred in the environment. This period, extending into the Silurian, is called the eophytic (proterophytic) or algal period.
An important plant developmental step occurred during the Silurian Period, where colonisation of land happened from the eophytic to the palaeophytic period. The first land plants possessed cells with large vacuoles, and some were able to develop stomata and other supporting tissues. These autotrophic plants lived together with fungi and bacteria (primary decomposers) communities and formed the first biocoenoses. From that period, plant diversity continuously increased through evolution.
Animal and plant communities evolved in unison (Table 17.1, Fig. 17.3). Psilophytes (archetypal ferns) are regarded as precursors of the Pteridophyta, to which Filicinae, Equisetinae and Lycopodiinae belong. Analysis of deposits of bituminous coal allow exact reconstruction of this vegetation, including indications of symbiotic interactions (mycorrhiza). At that time Bryophyta (mosses) split off and remained at this developmental stage. Other groups in the warm and humid climate of the Carboniferous formed luxuriant forests with tree-like Equisetinae, Lycopodiinae and Cordiates, which would later go extinct.
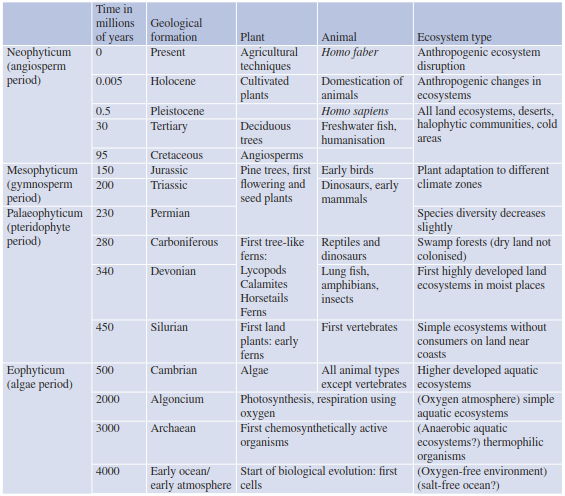
Table 17.1. Putative evolution of life forms and ecosystems during Earth’s history (after Kreeb 1983)
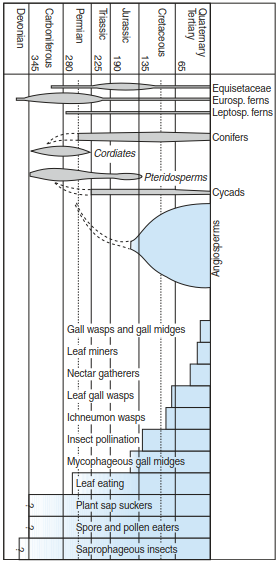
Fig. 17.3. Age and development of important groups of vascular plants and of phyto- and entomophageous insects. First occurrence of insect pollination and herbivory is also shown. (after Zwölfer 1978)
During the transition from the Carboniferous to the Permian Period, climatic conditions became much dryer. Many species became maladapted, which led to extinctions as many plant species were unable to adjust their water retention capabilities. Thus, the transition from the palaeophytic to the mesophytic is highlighted by a significant decrease in plant diversity. Colossal shifts in landmasses occurred at the beginning of the Triassic, when the Tethys Ocean separated from the eastern part of Pangaea. During the Jurassic, the North Atlantic developed, while the southern Atlantic formed during the Cretaceous Period. In the early Tertiary Period, Antarctica and Australia broke off as well, and it took up until the Pliocene to arrive at the present position of the continents we see today.
Up to the Cretaceous Period, global flora was generally similar in diversity. The oceans at this point were still too small to be considered barriers to the exchange of flora. The history of vegetation up to the mesophytic is also called the period of the gymnosperms. Following the extinction of the larger club mosses and horsetails, gymnosperms, particularly conifers, were able to expand their distributions. In the northern regions the first representatives of the Pinaceae and the genus Juniperus are found. Cupressaceae occurred worldwide. The boundary between the Jurassic and Cretaceous Periods also divides the mesophytic from the neophytic and the gymnosperms from the angiosperms. The first angiosperms occurred at the end of the Jurassic Period.
In 25 million years of the Cretaceous Period, flowering plants developed rapidly and outcompeted many of the gymnosperms, which up until then had dominated. Almost at a snap of the finger all the main angiosperm groups developed, and all suitable habitats were colonised. Intercontinental flora exchange was still possible and helped facilitate the rapid rates of colonisation. A stronger floristic separation occurred during the Upper Cretaceous Period. The so-called plant kingdoms developed and created the three floristic realms in the Southern Hemisphere, Antarktis, Australis and Capensis, two tropical equatorial floristic realms, Neotropics and Palaeotropics, and the Holarctic, the sole Northern Hemisphere floristic realm.
Zwolfer (1978) gave the first overview of geological occurrences of vascular plants as well as phytophageous and entomophageous insects, which addressed the development of communities. It was reported that both pollen and dead plant material were eaten by arthropods as early as the Devonian Period, and in the Permian leaves were consumed. In the Triassic, insect pollination of flowers occurred, and there were entomopha- geous parasites and plant galls. From the Tertiary onwards, all present-day phytophages are represented: miners, gall flies and gall gnats (Fig. 17.3).
Mutual adaptations of floristic and faunistic partners (vertebrates as well as invertebrates) result when selective pressures are linked in development. Plants developed thorns, spines or chemical defence substances, whereas animals adapted through changes in their mouth parts or developing resistance against plant toxins. Some flowering plants were able to protect themselves against herbivory, for example, Gentianaceae (defence by indoalkaloids) or Solanaceae (defence by tropanal- kaloids); however, again this led to the development in some animals of an ability to adapt to some of the most powerful plant toxins. In some cases animals made use of these substances for their own defence (Sect. 19.4, Chap. 19).
Most groups of flowering plants were present at the beginning of the Tertiary Period. The neophytic or angiosperm period is the current period. Among flowering plants, specialists are resilient in the most stressful environmental conditions (e.g. desert, highly saline soils, bare rocks). Along with the formation of mountain ranges, young mountain floras developed (oreophytes, sometimes closely related to flora in plains). Often individual altitudinal steps are characterised by vicariad species and genera. An example of altitudinal vicaria- tion in the Alps is Anthoxanthum odoratum for grasslands at lower altitudes and Anthoxanthum alpinum at higher ones. Another example of ecological vicariation is the alpine Rhododendron species R. ferrugineum, which grows on siliceous rock, and R. hirsutum, which grows at the same altitude but on calcareous rock.
Holarctic flora has been found in Palaeozoic to Eocene lignite deposits in Spitsbergen, where not only the remains of plants growing in today’s Central Europe under conditions of a temperate climate were found (e.g. Acer, Betula, Fagus, Quercus, Pinus and Picea) but also those that now grow only in humid, subtropical regions (e.g. Taxodium, Magnolia, Liriodendron). Plants found in Greenland represent the so-called arctotertiary flora, the basis of the Holarctic flora (e.g. Aesculus, Castanea, Platanus, Vitis). From the Eocene “flora from the Geiseltal” near Merseburg, Germany, plant lists were compiled containing tropical plant families (e.g. families Annonaceae, Pandanaceae, Sterculiaceae). The question thus arises as to how tropical genera can grow in areas currently characterised by cold-temperate climatic conditions. Plate tectonics is the justifiable answer, where Europe had only taken up its current position in the Northern Hemisphere during the Tertiary Period.
In plant remains found from the Pliocene in Central Europe, tropical species are absent, but present-day genera (Fagus, Quercus) indicate a worldwide progressive cooling. In Europe, climate and vegetation zones shifted to the south. The development of new barriers began to become an issue, where the west-east extension of the Alps and the Mediterranean Sea made it difficult for certain species adapted to a warm climate to survive, where some species went extinct and others were able to survive in local refuges or simply migrated to more favourable regions.
The relative uniformity of Pliocene flora in the Holarctic was lost. Large distribution ranges were separated into several spatially limited regions, the large Tertiary disjunctions (different separated parts of the former area of distribution, e.g. Sequoia and Metasequoia). For other species, for example, of the genera Styrax, Platanus, Melia and Castanea, a few moist warm sites became important refuges. The development of Mediterranean sclerophyllic vegetation is often connected to the general cooling of global climate in the Tertiary Period. Cooling in the Pliocene may be regarded as a precursor of the multiple quaternary climate changes, especially when compared with those occurring in the Pleistocene and Holocene.
Date added: 2026-04-26; views: 160;