Late and Postglacial Climate and Vegetation History
The vegetation history up to the Pleistocene has shown two main aspects:
- The enormous evolution of species and plant communities dependent on astronomic, tectonic and climatic events in biocoenoses.
- A distinct spatial separation of large parts of the Earth’s crust occurred, with orogenesis reaching the actual situation.
Now, in smaller areas, changes of the floristic characteristics due to small-scale differences in the site conditions are more important. Following the Holocene Period, the continents had almost reached their current position. The result was an irregular pattern with approximately 71% of the globe covered by water and the remaining 29% constituting continental land masses. The latter are unequally distributed with a large maximum in the Northern Hemisphere. Furthermore, large mountain ranges formed in Eurasia, extending mainly from west to east and north to south in the Americas. These main features have a determinant role in regional, continental and global climate, and, thus, impact the distribution and dynamics of vegetation.
Because of the outstanding example of the north-south extension of the Cordilleras, the different atmospheric conditions in the Americas, coupled with the regional pattern of glaciers and ice sheets, together form a developmental process different from what has occurred in Europe. Plant migration following the climatic changes was easier because difficult-to-cross west-east barriers like the European alpine mountain chains are missing. We concentrate here on the development in Eurasia and Africa.
The history of vegetation during the Holocene in Europe has been documented by pollen analysis and radiocarbon dating (Table 17.2). Almost all plant species found using these techniques have survived to the present day. Today, comprehensive data sets are available that are directly or indirectly linked to the vegetation development in postglacial and Holocene times. Not all historical analysis is done with pollen and radiocarbon dating; other techniques include macrofossils, tree-line changes (Fig. 17.5) and tree rings.
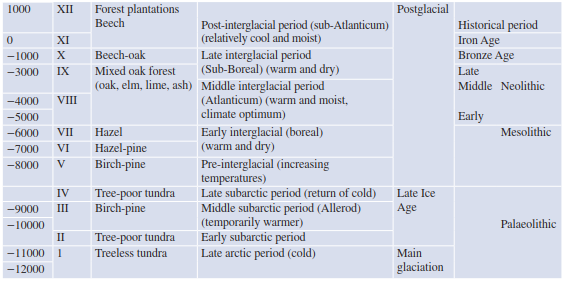
Table 17.2. Vegetation and climatic elements of late- and postglacial times. Columns for time, pollination zone I–XII (from Overbeck 1975), vegetation period, climate history and human prehistory (after Walter and Straka 1970)
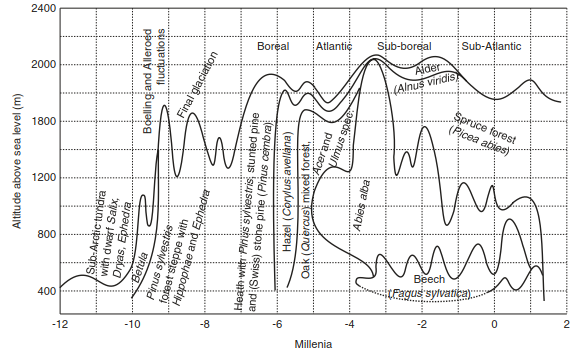
Fig. 17.5. Development of altitudinal zones of vegetation in northern Swiss Alps since maximum of Würm glaciation. (after Strasburger and Sitte 1998)
Various waterborne traces have also been used, where the measurements of inorganic compounds within glaciers, ice sheets, eustatic terraces and lake levels can be taken. Furthermore, changes in orbital axes and solar radiation can also be used to draw inferences from the patterns and the likelihood of the presence of certain species in the past. Finally, for the last few centuries anthropogenic greenhouse gases and remarkable land cover changes have contributed directly or indirectly to more recent vegetation changes.
On the basis of the various sources of data, some models have been developed and have proposed findings that suggest detailed accounts of climate fluctuations leading to reactions of plant communities. Controversy still surrounds the causality of these findings, which continue to be discussed (Gallimore et al. 2005; Holzhauser et al. 2005). The difficulty in attributing the causes to one factor or a number of factors lies in the fact that there are many aspects that contribute in combination with a high order of complexity in terms of the processes and how they can change with climate. The differences are often due to the different data included in modelling that have been considered as decisive (e.g. atmosphere-ocean-land data, glacier or lake-level changes, solar activities or albedo changes).
So far, the results allow two general interpretations. The first involves researchers seeing a possibility or a necessity to put the results in a system of periods. This method has been traditionally done in pollen analysis, which was built from the diagrams of Firbas (1952 ff) and Overbeck (1975) and later improved by Holzhauser et al. (2005) and Joerin et al. (2006). The second interpretation involves incorporating variability in an attempt to discern general trends, for example, cooling or warming, under humid or arid conditions (Prentice and Jolly 2000; Wanner et al. 2008).
For a description of the late glacial time and the Holocene we have divided the following into three phases:
- Late- and postglacial times and early Holocene (18,000-9000 BP).
- Mid-Holocene (9000-6000 BP).
- Late-Holocene and preindustrial time (6000 BP-1700 AD).
In these three phases the development of the vegetation is closely connected with climatic changes. The influences of the late Holocene, where a remarkable human aspect begins to be identified on the vegetation, will be dealt with in the next subchapter.
In the late glacial period, also called the Dryas Period after the indicator species Dryas octopetala, a continental steppe dominated by scattered tree-free tundra islands was reconstructed after the results of pollen and macrofossil vegetation relics. All over the Northern Hemisphere, the cold, dry conditions led to different tundra and grassland types. Forest biomes were displaced southward, the boreal (i.e. taiga) biome became largely fragmented, while at the same time the moist tropical and temperate forests saw significant reductions. It is argued that during this period the arid Saharan mountains we know today may have been linked with the Mediterranean and tropical forest biomes and may have led to some of the discontinuous or fragmented range disjunctions, for example, Acacia spp. and Euphorbia balsamifera (Deil and Muller-Hohenstein 1984).
A noticeable warming of the climate occurred around 12,000 BP and was accompanied by the retreat of alpine and continental glaciers. As a result of this retreat of the glaciers, many species filled the newly created void of space. Warmdemanding species, in particular trees, were able to take advantage of this space in Central Europe. In Northern Europe, the forest biomes extended into the present-day tundras, while in the South, large parts of the Sahara were invaded by tropical grassland and dry shrubland.
This did not occur continuously. The migration of the tree species back to the earlier positions was reconstructed by comparing pollen profiles, so-called iso-pollen maps. This dynamic occurred in different ways and at different periods, according to the requirements of species for the site, the speed of migration and the location of the refuges during cold periods. Spruce (Picea) immigrated from an easterly direction, following the “northern path” from the southern Urals. Fir trees (Abies) and almost all deciduous trees (e.g. Fagus) originated from the Mediterranean areas and reached Central Europe via the Burgundian Gate or the Pannonian Basin. A pollen diagram from the Lutter Sea in northern Germany, representative of Central Europe, shows a detailed picture of the development of the temperate deciduous broadleaf forest biome from the late glacial period on (Figs. 17.6 and 17.7).
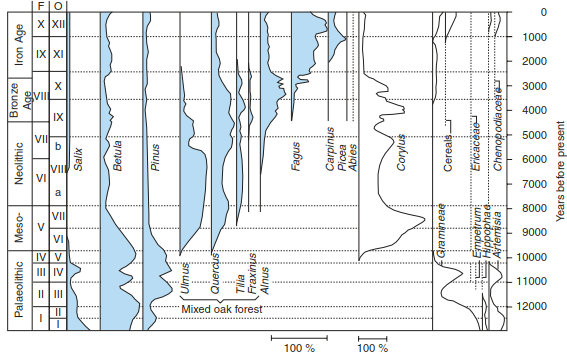
Fig. 17.6. Pollen diagram from Luttersee. The lake is situated near Göttingen, northern Germany, 160 m a.s.l.). Scale: sum of tree pollen 100%, without hazel (Corylus). (after Walter 1986)

Fig. 17.7. Undisturbed and disturbed raised bogs. a Part of the almost natural “Pillermoor” (1600 m a.s.l.), which is a protected area in the lower Pitztal (Tyrol, Austria). b Part of same raised bog where peat was removed more than 30 years ago. Pollen analysis from this bog shows the postglacial development of vegetation of the montane region in Tyrol. (Photos: K. Müller-Hohenstein)
During the following 6000 years, the temperatures remained high with a number of distinct fluctuations. Consequently, the vegetation cover also changed; in the warmest periods (e.g. Allerod period, Atlantic period), sparse spruce and birch forests and willow shrubs were able to exist. The steppes advanced from the east under the still extremely continental conditions. The temperate forest was able to extend; birch was methodically replaced by hazel and other deciduous trees. Human populations in the Mesolithic (about 8000 BP) were surrounded by species-rich forest communities. Remnants of tundra occupied the increasingly melting edges of the continental ice sheet, and steppes were pushed by invading forests towards the south and south-east.
This development is also confirmed by results of tree-line studies. The position of the alpine tree line in the last 12,000 years was approximately 1200 m lower in the last glacial period compared with present day conditions, outlined by the remarkable warming over this period (Fig. 17.5). The variation of the tree line, as well as a recently published temperature reconstruction, outlines similar temperature fluctuations following this temperature increase in the middle-late glacial period (Marcott et al. 2013). Throughout the early Holocene the temperatures stayed—with some regional variations—at this level. In the European Alps the tree line was about 400 m higher than today.
The warmest climatic conditions, and the corresponding development of vegetation, were experienced by the early Neolithic people in the Atlantic period about 6000-7000 BP. In Northern Europe, during this time of the “climatic optimum”, mean annual temperatures were 2-3 K higher than present temperatures. Mixed oak forests with deciduous trees requiring warm climatic and nutrient-rich edaphic conditions were dominant. In the montane areas, fir dominated in the western Alps, spruce in the eastern Alps and pines in the eastern part of Central Europe on nutrientpoor sites. Thermophilic species still occurred at dry sites with grassland vegetation.
Glacial relicts from the cold periods are still maintained in present-day vegetation in isolated sites, for example, Betula nana in the foothills of the Alps in peat bogs. Similarly, species from the warmest periods were also able to persist on limestone outcrops. These xerotherm relicts, sub-Mediterranean species (Quercus pubescens) and those from the south Siberian and Pontic areas (Stipa spp.) were able to survive after the onset of cooling in the Sub-Boreal forest. To ensure their survival, both of these relict species required specific microclimatic or edaphic conditions. For xerotherm species, extremely shallow and southern facing, exposed sites, which inhibit the growth of trees, are needed.
Beech only became dominant in the forests of Central Europe in the Atlantic period and took over the role of hornbeam (Carpinus betulus), which dominated in the interglacial period and was now predominant further to the east. Together with spruce and fir, beech forms forests up to the montane belt. Even though beech has dominated the forests of Central Europe for only a relatively short time period (30-50 tree generations), beech-dominated forests became established everywhere, up until increased human intervention became more intense (Hasel and Schwartz 2006).
Two main causes were responsible for further changes of the vegetation cover in the following millennia:
- a continuous decline in temperatures and.
- growing human influence (Sect. 17.2.4).
The climatic cooling (between 0.1 and 0.3 K over 1000 years) was the most important reason for large-scale plant migrations. Species that required higher temperatures migrated back into warmer regions and outcompeted the cold-period vegetation, except for a few relict species. Their expansion partly followed the edges of the ice sheet, and thus many closed areas became divided, and disjunctions were formed. Some of these disjunctions are concentrated on the Southern European mountains (e.g. species of the genera Primula and Saxifraga), while others form the arcto-alpine disjunctions with interspersed occurrence on the highest parts of the mountains of central Germany (e.g. Nigritella nigra agg.). Supplemental examples of disjunction distributions include the Betula nana and Salix herbacea and the circumpolar Loiseleuria procumbens and Eriophorum scheuchzeri in North America.
A reconstruction of the changes in global temperatures shows a constant decline in temperature, which ended only in the nineteenth century (Marcott et al. 2013). Especially thanks to a comprehensive palaeo Vegetation Mapping Project (BIOME 6000; Prentice and Jolly 2000), our knowledge about vegetation development from the mid-Holocene on has been considerably deepened. The reconstruction of the vegetation cover was made possible through the use of extensive pollen and macrofossil records. Vegetation was classified on the basis of plant functional types (PFTs), biomes, and mega- biomes with worldwide contributions to this programme (Harrison and Prentice 2003). Because of high summer insolation and warmer temperatures, plant species with high heat demand experience very advantageous conditions. Therefore, the vegetation distribution compared to present- day conditions, and not including anthropogenic influences, has led to a significant northward shifting of boreal and temperate species in the Northern Hemisphere. Although variability exists and depends on the region, all distributions were more than 100 km further north compared to today. Warmer winter temperatures also favoured a change from cold-temperate mixed forests to temperate deciduous forests. The Mediterranean subtropics, however, were covered with temperate grassland and xerophytic shrubland.
Only in recent decades have new data been collected in connection with the history and climate of the arid African areas (Holzhauser et al. 2005; Knorr and Schnitzler 2006). According to these findings, the long-established views of a synchronous sequence of cold and warm periods in higher latitudes (e.g. Central Europe) and dry and humid ones in lower latitudes (e.g. Northern Africa) must be corrected.
During a long period (approximately between 18,000 and 6000 BP), most of the Sahara was “green”, and warm-temperate mixed forests approached from the north as well as semi-evergreen broadleaf forests from the south (according to the original biome classification by Prentice and Jolly 2000). Mountains in the Sahara possessed a strongly Mediterranean vegetation. These conditions persisted up until the Atlantic period with semi- arid conditions and summer rains in the south and winter rains in the north, ultimately led to the Sahara being narrowed to a smaller region. Neumann (1988) assumes on the basis of charcoal analyses that Sahelian vegetation was approximately 600 km further north and dominated by such species as Balanites aegyptiaca, Acacia albida and Cadaba farinosa, which made up a sparse dry woodland, where grasslands advanced from 18.75° to 22.5°N.
Other vegetation shifts were also identified, although smaller in size: the south-west shift of Mediterranean species is also assumed to have happened due to a positive precipitation-vegetation feedback, based on changes in albedo. It wasn’t until about 6000 BP that everywhere in the Northern Hemisphere started cooling and experienced many more changes in vegetation. After weather conditions around 8000 BP began to cease, Neolithic man in the Sahara experienced increasing drought from 5000 years ago until now (Fig. 17.8).


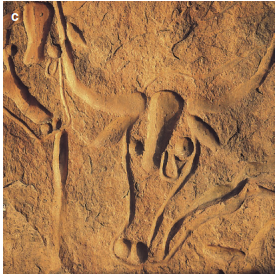
Fig. 17.8. Rock paintings and engravings indicate once more humid climatic conditions in central Saharan mountains. Pictures of hunters a and large grazing herds b near Sefar (Tassili), southern Algeria, are nearly 8000 years old. The “crying cows from Terarart” c near Djanet are about 5000 years old and indicate a gradual increase in aridity. (Photos: K. Müller-Hohenstein)
The tropical savanna reached its actual position again around 3300 BP. For the Sahara, even today, two relatively stable and interchanging vegetation phases are assumed, a “green” Sahara and a “red” Sahara that is almost bare of vegetation (Claussen 2001). The actual human-influenced expansion of desert conditions in all sub-Saharan regions leads to increasing losses of both plants and animals, even if the situation for the flora is less dramatic (Le Houerou 1997).
There is sufficient evidence from glacier- and lake-level studies in the Swiss Alps with high- resolution data that natural driving forces still contribute to climate change (Holzhauser et al. 2005). Also, the results of tree-line studies from Russia underline a continuous cooling, at least more or less synchronous for the continents of the Northern Hemisphere (MacDonald et al. 2000a, b). Only in the last 300 years, despite the input of volcanic aerosols and a certain variability in solar insolation (both of which initiate cooling effects), has it been exceeded by the warming effect of the input of excessive greenhouse gases and land-use changes (shifting of forest to agricultural lands), which has ultimately led to a reverse forcing of the biophysical drivers for the continuous cooling seen in many regions. The main driving force for all these changes in the vegetation cover is no longer exclusively a change in climate; we must now consider anthropogenic effects in all biomes.
Date added: 2026-04-26; views: 159;