Influential environmental factors. Water balance
Introduction. As was pointed out in Chapter 1, there are indications that variations in the abiotic environments of inland waters strongly influence molecular and morphological evolution by altering mutation rates and exposing cryptic variation. Extreme environments, such as temporary waters, are likely to be particularly influential, and have been hypothesized as foci of biological diversification. Major parameters in the physical and chemical environments of temporary freshwater ponds and streams are summarized in Figure 3.1. Some are dealt with below under separate headings, but others, such as light and pH, are interactive and are discussed in multiple contexts. Biological influences are introduced at the end of the chapter.
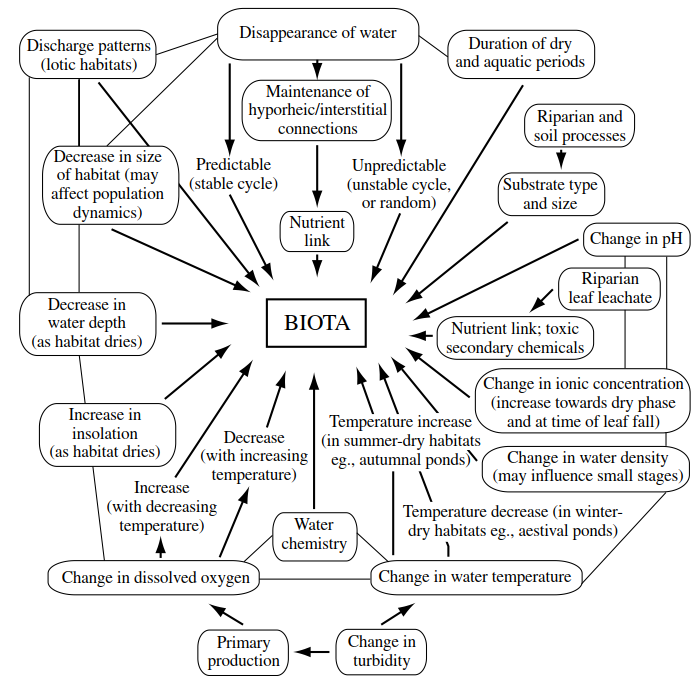
Figure 3.1. Summary of the major physical and chemical factors that influence the biotas of temporary freshwaters (from Williams 1996; an arrow indicates that the ‘boxed’ factor has been shown, or is likely, to have an effect on organisms—sometimes indirectly through another boxed factor; interactions between factors are shown by the lines joining the factor boxes)
Water balance. Of fundamental importance to the existence of any body of water is the result of the balance between water gain and loss. For a permanent stream or pond, water input equals water loss. However, as noted in Chapter 2, there are several sources of input (surface run-off, groundwater flow, precipitation, etc.) and several forms of output (loss downstream, absorption by the soil, evaporation, uptake by plant roots, etc.), the magnitudes of which vary in space and time causing water levels to fluctuate. Waterbodies in which input and output rates are highly variable are frequently temporary.
The exact length of the aquatic phase (the hydroperiod) varies according to both geographic location and local hydrological conditions, and is also related to water depth and surface area. For example, in the tropics, temporary pools and streams contain water immediately after the monsoons but may soon lose water due to evaporation by the sun; in areas of extended high rainfall, the life of the water body will likely be extended until the dry season. At high latitudes, although evaporation may not be as severe, a pond may dry up for a short period in late summer but may also 'lose' its water in winter due to freezing solidly to its bed. By way of illustration, Figure 3.2 shows the water level and some associated variables in Sunfish Pond, Canada.
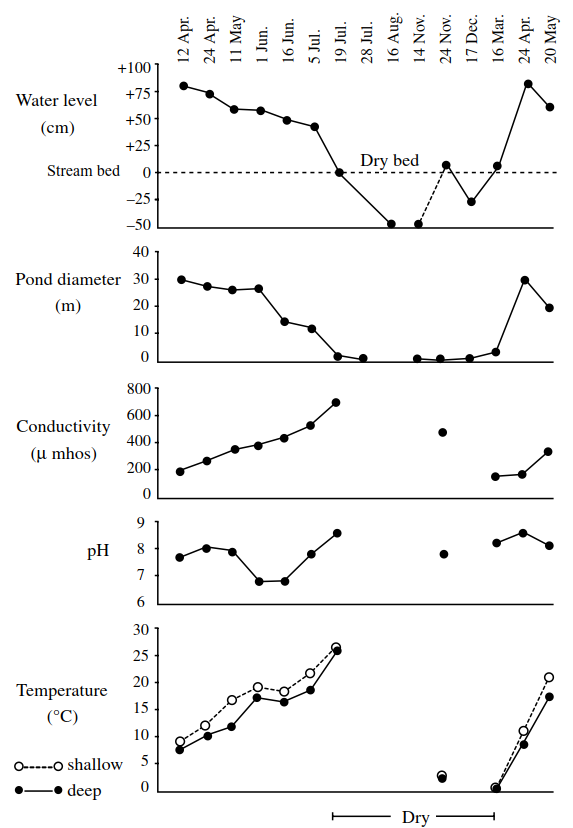
Figure 3.2. Characteristics of the hydroperiod and some associated parameters in Sunfish Pond, an intermittent, vernal pond
This temperate zone, intermittent pond receives most of its water in spring from snow-melt. At this time, loss to surrounding soil is negligible as the pond margins are either saturated or still frozen. Evaporation is also small because of diminished solar radiation. As time passes, solar radiation intensifies, air and water temperatures rise and evaporation increases. Simultaneously, the groundwater table drops and the water level in the pond decreases. Eventually, in mid-summer, the pond becomes dry. Occasionally, very heavy summer rainfall may cause the pond to fill temporarily (a few days), but usually the pond remains dry until the following spring. In years with low snowfall or an early, warm spring, the hydroperiod may be shortened. Conversely, in years with high snowfall or a late, cold spring, it will be extended. Wiggins et я/. (1980) termed this kind of pond 'vernal', to distinguish it from intermittent 'autumnal' ponds which flood in the autumn and remain wet until the following summer.
Temporary streams, as noted, experience several distinct phases in their hydroperiod: surface flow connected to the GWT, reduced flow disconnected from the GWT, isolated pools (detention storage), and dry bed (Figure 2.4). During the latter, heavy rains may again allow brief surface flow. A number of discharge-related factors, shown in Figure 3.1 (e.g. temporal pattern, maxima-minima, maintenance of connection with the hyporheic zone), are likely to influence the biota both directly and indirectly; the latter through correlation with several of the other physicochemical factors (such as water depth and dissolved oxygen content) shown in the model.
The pattern of water loss is important particularly in terms of whether it is predictable (i.e. part of a stable cycle) or unpredictable (i.e. random, or linked to vagaries of local climate). One might assume that adaptations to deal with predictable disappearance of the water (intermittent waters) evolve more readily than those required to deal with random disappearance (episodic waters). However, a strategy such as 'bet-hedging' (Stearns 1976; Baird et al. 1987; see Chapter 5) would seem suited to both situations— although it might optimize rather than maximize reproductive effort in episodic waters.
Comparison of the invertebrates living in three water-filled ditches in southern Ontario (Table 3.1) indicates a decrease in species richness with decreasing length of the hydroperiod. A similar relationship between species richness and flow duration has been shown for some streams in Australia (Boulton and Suter 1986). However, in a study of six, small upland streams in Alabama, Feminella (1996) found that invertebrate assemblages differed only slightly, despite large differences in flow permanence: 75% of species (primarily insects) were common to all of the streams, or showed no relationship to permanence; only 7% were exclusive to the normally intermittent streams. Indeed, inter-year differences in assemblages within single streams were as great as within-year differences among streams of contrasting permanence.
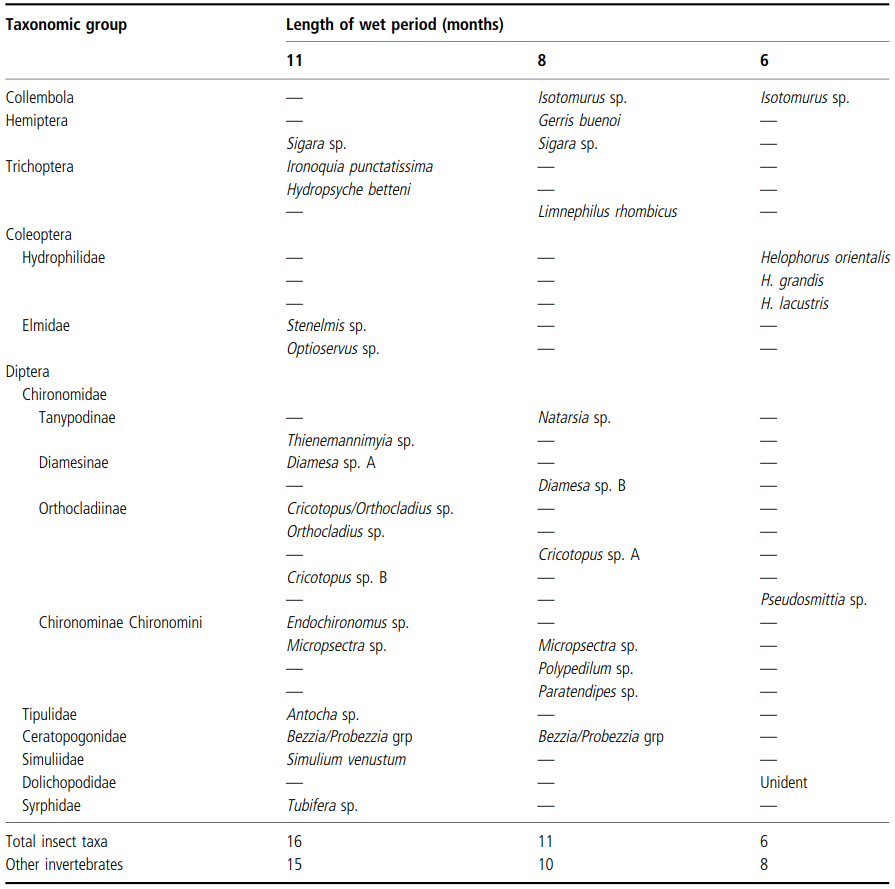
Table 3.1. Comparison of the insect and non-insect faunas of three Ontario water-filled ditches with different lengths of hydroperiod, shown as the number of months in the year in which water is present. Absence of a particular taxon is shown by a dash
In a comparison of crustacean diversity and hydroperiod in standing waters in North America (Table 3.2), Williams (2002) concluded that:
(1) maximum taxon richness occurred in habitats with a hydroperiod of between 150 and 250 days;
(2) some additional taxa occurred in habitats with hydroperiods of between 70 and 150 days, but no additions (to the genus level) occurred in habitats wet for less than 70 days; and (3) richness of the classic, three, large temporary water branchiopod groups (Anostraca, Notostraca, and Conchostraca) fell dramatically in habitats containing water for more than 250 days per year.

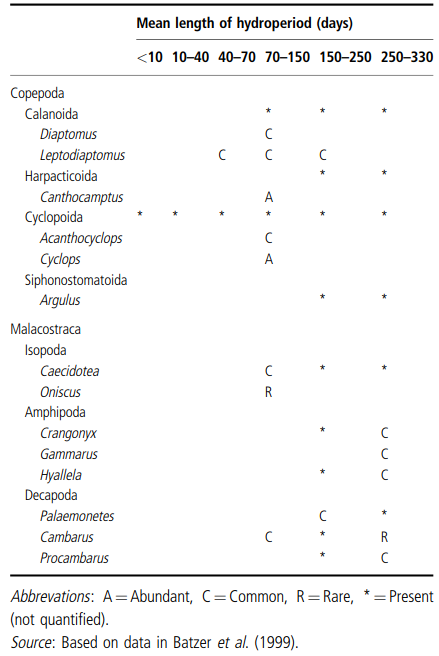
Table 3.2. Occurrence of crustaceans in North American temporary waters in relation to length of the hydroperiod
In floodplain ponds of the Mississippi River, species richness and abundance of predatory invertebrates increased with increasing hydroperiod, whereas overall invertebrate abundance and richness decreased (Corti et al. 1997). Boix et al. (2001) similarly found a positive relationship between hydroperiod duration (and also flooded surface area) and species richness in Espolla Pond, Spain. Hershey et al. (1999) found that, in the prairie wetlands of Minnesota, for some groups (especially insects) species counts decreased with shortening hydroperiod, however, for others (e.g. molluscs) there was no relationship. In ponds in central Italy, Bazzanti et al. (1997) have shown that in those with a long hydroperiod (but also lower oxygen content), the chironomid assemblage was dominated by members of the Chironominae and Tanypodinae. A pond with a shorter hydroperiod, but also higher oxygen, was dominated by Orthocladiinae. Suter et al. (1994) found that chironomids responded to flooding and drying events in the floodplain wetlands of the Lower Murray River in South Australia. Notably, species of Chironomus responded rapidly to flooding, and species of Procladius (Tanypodinae) became more abundant as drying progressed. However, these authors cautioned that while, for some species, the responses were consistent across several locations, for others they were not. Timing of flooding, salinity variations, and colonization sources were all thought to have some influence.
Temporary coastal rockpools in Jamaica exhibited low variability in community structure, and were dominated by 'weedy' species with good powers of dispersal and colonization (Therriault and Kolasa 2001). Schneider (1999) concluded that the invertebrate communities of short hydroperiod, snowmelt ponds in Wisconsin were structured primarily by species adaptations to the threat of drying (i.e. to abiotic factors). On the other hand, the communities in ponds with longer hydroperiods were structured more by biotic interactions, particularly predation and competition. Greater consideration of such r- and K-selected species will be left until Chapter 5.
Working on the largely crustacean-dominated pans of South Africa, Meintjes (1996) found that larger pans supported more species than smaller pans. Decreasing water volume in tree hole habitats has been shown to induce metamorphosis at smaller sizes in females of the mosquito Aedes triseriatus, indicating that larvae can detect changes in the hydroperiod and take appropriate steps to escape a deteriorating environment ( Juliano and Stoffregen 1994). As to the mechanism involved, a laboratory simulation experiment using the frog Rana temporaria demonstrated that tadpoles were able to respond to pond drying by speeding up their development through behavioural mediation, irrespective of any change in water temperature (Laurila and Kujasalo 1999).
Date added: 2026-07-14; views: 3;