Climate, seasonality, and habitat persistence
Regional climate is clearly a major factor in determining the nature of the local precipitation/ evaporation balance, and hence the propensity for temporary waters to form. Rainfall frequency, amount and pattern, are all important, as are the various mechanisms of water loss. As already noted, this balance is also influenced by nearsurface geomorphology (basin/channel availability), vegetation (density and type), and land use. The various permutations of these factors together with climate produce temporary water habitats on all continents, whether they be episodic streams in arid regions, or intermittent, vernal or autumnal ponds in the temperate zone.
The pattern of water availability is particularly important as it often characterizes the type of habitat and the physicochemical signature that its biota will have to endure. For example, in India and much of the Oriental tropics there are the annual monsoons. Here, warm and moist air from the ocean is blown north where it encounters either high mountain ranges or cool, dry air moving south. The result is heavy rainfall from June to September resulting in the establishment of a host of temporary water habitats, ranging from extensive wetlands, to floodplain lakes, to rain- filled rockpools. The latter are likely to be characterized by water that is low in nutrients and electrolytes, reflecting a direct rainwater origin; the former, in contrast, are more likely to have turbid, more nutrient-rich water resulting from overspill from adjacent, swollen rivers.
Seasonality is also a strong feature of temporary ponds and streams formed by snow melt at higher latitudes, and of early summer glacier melt at high altitudes. The influence of growth season on various phytotelmata has already been discussed. Even the availability of man-made habitats may reflect seasonal activities within cultures, for example, the flooding patterns of irrigation canals based on crop growth, and the summer rejuvenation of ornamental water fountains.
An aspect of climate that will likely impact temporary waters markedly in the future is global climate change. The earth's climate is being warmed, as a result of the accumulation of greenhouse gases, such that within 100 years not only will the earth be warmer than it has been for a million years, but the rate of increase will have been greater than any on record (Schneider 1989). A range of climate-related changes are expected to accompany a doubling of atmospheric carbon dioxide, including melting of the polar ice sheets and permafrost, and rising sea level (Poiani and Johnson 1991). The mean summer air temperature of the Great Plains of North America, as an example, are predicted to rise by 1-2°C, with an even greater rise in winter (3-4°C). Although less certain, models foresee increases in mean global precipitation of 3-11%, but accompanied by faster return to the atmosphere due to the warmer temperatures (Bradley et al. 1987).
Ironically, continental interiors may become more arid as a result of a shift in current mid-latitude rain belts. All of these changes will impact the numbers, types, and properties of temporary water habitats. To appreciate this, Figure 2.8 illustrates the hydrological budget of a prairie wetland pond. In reality, this pond was classed as being 'semipermanent' (i.e. holding water throughout the growing season during most years) (Poiani and Johnson 1991).
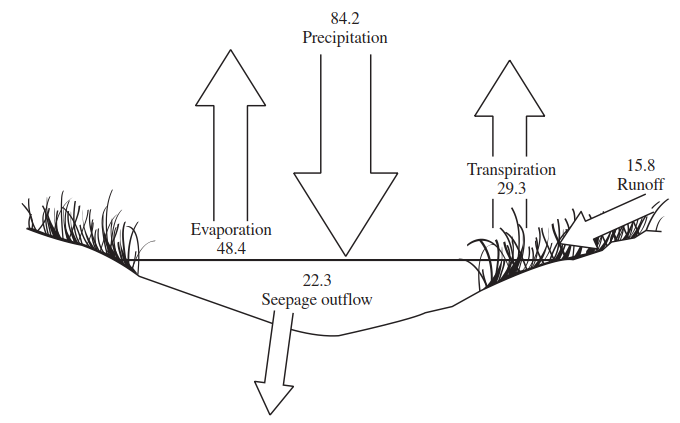
Figure 2.8. Hydrological budget for a prairie wetland pond (water loss/gain units are expressed in percentages; modified from Poiani and Carter 1991)
This type of pond thus sits on a hydrological knife-edge between being permanent or temporary. It is easy to see how it's balance, and that of many other types of temporary waters, could be tipped by changes in local climate: for example, more precipitation and it will never dry out; less precipitation, accompanied by an increase in seepage outflow by virtue of a lowered GWT, and it will become very short-lived. Further, Manabe and Wetherald (1986) predicted a 30-50% decrease in the summer moisture levels of Great Plains' soils, which could adversely affect the survival of the dormant stages of pond bed biota (see Chapter 5). In contrast, rising sea levels would flood many coastal temporary waterbodies, rendering them permanent with the loss of drought-adapted species.
In another modelling exercise, Larson (1995) looked at the effects of climate-related variables on the number of wetland basins occurring in the northern prairies. Her models which included information on spring and autumn temperatures, annual precipitation, the previous year's basin count, and the previous autumn's precipitation— accounted for 63-65% of the variation in the number of wet basins. She also demonstrated that wetlands in more wooded areas ('parkland') were more vulnerable to increased temperatures than those in grasslands.
Some of the predicted, as well as the more complex, effects of global warming are already being seen in North American lakes (Schindler et al. 1996). For example, Yan et al. (1996) observed that in a boreal lake that experienced a drought- related drop in water level the increased exposure caused a re-oxidation of sediment sulphur, which resulted in a re-mobilization of acid into the lake water. This produced a decrease in dissolved organic carbon concentrations that was sufficient to increase, by three times, the depth to which ultraviolet radiation could penetrate.
Another climatic phenomenon that is likely to significantly affect temporary water habitats is El Niho. Under normal conditions, easterly winds blow across the equatorial Pacific pushing sea surface water westwards. The water thus displaced from the west coast of South America is replaced by upwelling of cold, deep-ocean water that lowers sea surface temperatures in the eastern Pacific. This promotes a cool, dense air mass that fails to rise sufficiently high to form rain clouds, thus creating a high pressure system, with cool dry conditions. Low pressure systems, producing monsoon rains, are therefore confined to the warmer, western Pacific. During an El Niho event, winds shift so that surface water is no longer pushed away from the west coast of South America. Deep-water upwelling diminishes, or ceases, and seas surface water and air temperatures rise in the eastern Pacific, producing low atmospheric pressures. This results in the monsoon rain belt extending eastwards into the central Pacific, creating drought in the western Pacific and flooding in the east. In addition to these regional changes, El Niho produces changes in the circulation of the upper atmosphere as dense, tropical rain clouds rise, altering the positions of both global rain belts and jet streams. The net effect is to produce unseasonal weather in many parts of the world, including shifts in the frequency and duration of floods and droughts
Habitat persistence is another important aspect of temporary waters, although its strict definition 'being permanent' (Oxford Concise Dictionary), seems at odds with the basic nature of temporary waters. But it is necessary to distinguish between the persistence of an individual temporary water body and the persistence of that type of temporary water body within a given locality; the former may be short-lived (e.g. a rain puddle created in a tyre track), whereas the latter may persist over a much longer timespan (e.g. pools on the floodplain of a major river). Such floodplain pools may be shortlived, individually, but, collectively, they allow establishment of metapopulations of suitably adapted temporary water biota on the floodplain. Indeed, a hypothesis was aired in Chapter 1: that temporary waters represent very ancient habitats that may have played a significant role in the origins of life (W.D. Williams 1988).
Many equate habitat persistence/permanence with habitat stability, for example, the stability of ancient Lake Baikal is often cited as the main reason for the preservation of its endemic flora and fauna. In terms of stability, then, how do temporary waters compare with, for example, freshwater springs, which, with their classically constant discharge and uniform water temperatures, would seem to be at the opposite end of a stability/permanence axis? Although the field data are limited, some tentative comparisons can be made at the regional level. In southern Ontario, permanent cold-water springs are dominated by nemourid stoneflies, chironomids, caddisflies, mites, copepods, ostracods, and amphipods (Figure 2.9).
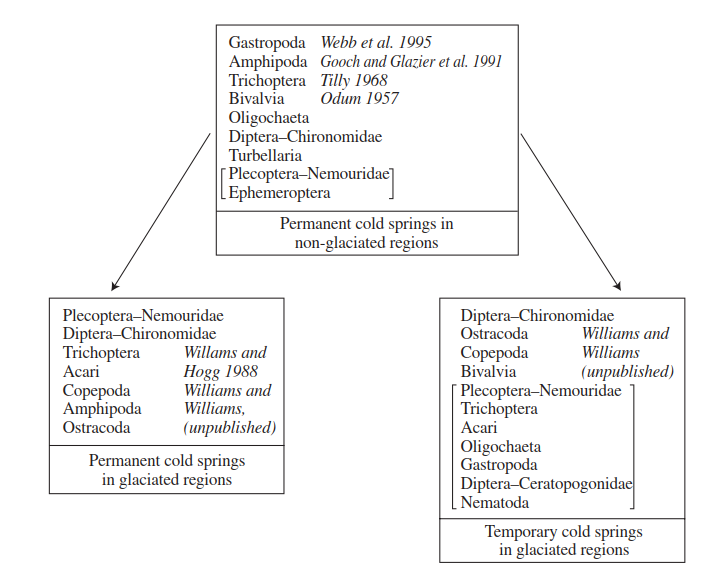
Figure 2.9. Comparison of the dominant invertebrate taxa known to occur in temporary and permanent, cold-water springs in southern Ontario, Canada, together with those found in permanent springs in non-glaciated regions. In each box, the taxa are ranked from top to bottom in the approximate order of numerical dominance seen in that spring type. Taxa enclosed within the square brackets are less common but usually present. It is possible that, due to non-standard collecting mesh size, the microcrustaceans may have been underrepresented in some of the studies from which the data were gathered
With the exception of the mites and chironomids (e.g. 20 species and 10 genera, respectively, present in Valley Spring, Ontario; Williams and Hogg 1988), diversity within these taxa is not high. For example, there are usually only one or two species of stonefly and amphipod per spring, although population densities can be quite high. Intermittently flowing, cold-water springs in southern Ontario have a fauna that is taxonomically similar to that found in permanent springs, however, there is a predominance of chironomids, ostracods, copepods and sphaeriid clams. Although moderate numbers of caddisfly larvae are sometimes present, they do not match the dense populations recorded from a temporary spring in California in which trichopterans dominated the fauna (Resh 1982).
Figure 2.9 also compares the faunas of these southern Ontario springs with those from adjacent regions of eastern North America that were largely unaffected by recent glacial activity (see Matthews 1979). The latter, geologically more persistant habitats, are dominated, numerically, by gastropod and bivalve molluscs, triclads, amphipods, oligo- chaetes, and trichopteran and chironomid larvae. Nymphs of nemourid stoneflies and mayflies are present often, also. The diversity of species within some of these taxa may be very high, for example, 17 species of Oligochaeta in Old Driver Spring, southern Illinois (Webb et al. 1995). The insect/non-insect dominance seen between permanent, cold-water springs in Ontario and those to the south may well be related to respective glacial histories, with the more vagile insects predominating in less persistant habitats in regions that have been subject to recent glacial activity.
Date added: 2026-07-14; views: 4;