The Osmotic and Ionic Component of Salt Stress
When a plant, be it salt tolerant or sensitive, is subjected to a sudden increase in the NaCl concentration in the medium or soil, a fast and a slow reaction can be differentiated (Fig. 7.30 and 7.31). Leaves of various cereals and dicots show an instantaneous halt of expansion. Because of its rapidity and the subsequent partial recovery, this fast response must be due to changes in cell water relations. Since the same phenomena can be triggered with KCl, mannitol or polyethylene glycol instead of NaCl, this response is not salt specific.
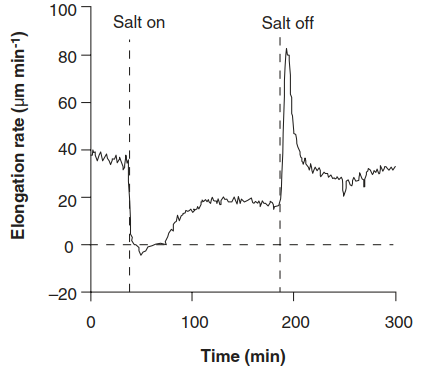
Fig. 7.30. Effects of changes in the salinity of the soil solution on the elongation rate of a barley leaf. A sudden increase in the salt concentration triggers an almost immediate drop in the growth rate, followed by a slower phase of partial recovery
For most plants, the threshold forthat response is around 40 mM NaCl, but it may be lower for particularly salt-sensitive species. Several minutes after the initial decline in leaf growth, a partial recovery takes place and a new steady growth rate is attained after approximately 30 min. The time required for recovery depends on the concentration of the salt solution. Removal of the salt results in an equally sudden overshoot followed by a fast return to the original growth rate.

Fig. 7.31. Short- and long-term responses of plants to salinity stress. The solid green line indicates the reaction of a plant to the two components of salt stress: an increase in osmolarity and toxicity of NaCl. The broken green line represents the reaction of a plant adapted to better tolerate osmotic stress, while the dashed red line indicates the reaction of a plant with higher tolerance of the ionic toxicity
Under long-term salinity stress, inhibition of shoot growth encompasses reduced leaf expansion, delayed formation of new leaves and delayed or even suspended bud break, resulting in fewer branches or lateral shoots. Contrary to the expectation, root growth in saline media is less affected than shoot growth. This might be due to the fact that a reduction in shoot growth also reduces the water consumption by the plant and mitigates the transpiration-induced increase in salinity in the rooted soil volume.
The Ionic Component of Salt Stress. Homeostatic ion concentrations in the cytosol of a non-stressed cell of a glycophyte—that is, a plant without adaptation to saline habitats—are 100-200 mM K+, 1-10 mM Na+ and Cl-, and 0.1-0.2 mM Ca2+. These concentrations are maintained in the plant cell mainly through ion transport energised by H+-ATPases and H+- pyrophosphatases in the plasma membrane and the tonoplast, respectively. In addition, channels and pumps are involved. The combined activities result in a membrane potential of around -150 mV at the plasma membrane and a tonoplast potential between +20 and +50 mV (the cytosolic side is always more negative). These membrane potentials are associated with differences in pH between the cytosol (pH 7.5) on the one hand and the apoplast and the vacuole on the other, whose pH values are around 2 units more acidic (Fig. 7.32).
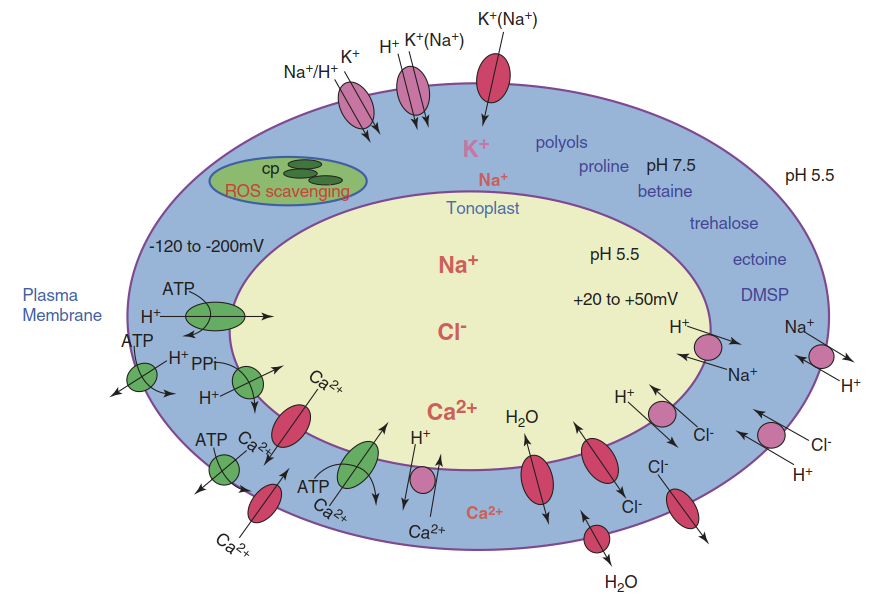
Fig. 7.32. Establishment of cellular homeostasis during salt stress acclimation, showing intracellular ion distribution, membrane potentials and pH values. Ions and osmolytes compartmentalised in the cytosol and vacuole, transport proteins involved in Na+ and Cl- homeostasis, and water channels are indicated. ATP- or pyrophosphate-driven pumps are indicated in green (note that for lack of space the products of ATP- or PPi-hydrolysis are not shown); channels gated by the membrane potential are indicated in red while proteins mediating secondary active transport are shown in purple
High salinity subjects this homeostasis to considerable strain (Fig. 7.31). In the majority of plant species grown under salinity, Na+ reaches a toxic concentration before Cl- does, and so plant chloride relations have attracted much less attention than the plant’s response to sodium (Munns and Tester 2008). Chloride is an essential micronutrient (e.g. for photosynthesis), acts as counter anion to stabilise membrane potentials and is involved in turgor and pH regulation. In the presence of high external salt concentrations, large intracellular Na+ pools build up, partly at the expense of the K+ pools. More than 50 enzymes are controlled or activated by K+, and Na+ cannot substitute for K+ in this role.
High concentrations of Na+ or high Na+ to K+ ratios therefore disturb or even disrupt various enzymatic processes. Photosynthesis and respiration are among the processes that are most sensitive to salt stress. Protein synthesis requires high concentrations of K+—for example, for the binding of transfer RNA (tRNA) to the ribosomes—and is thus also highly affected by high intracellular Na+. Finally, ion imbalances (e.g. in photosynthesis and respiration) and water shortage result in oxidative stress which, in combination with the aforementioned impairments, can easily result in cell and organ death.
Date added: 2025-02-01; views: 678;