General Vegetation Dynamics
Vegetation units are permanently subject to spatial and temporal changes. This concerns changes in species composition as well as the constituent life forms and alterations in vegetation structure. Each recording of vegetation is only a momentary snapshot of a complex dynamic process. The temporal dynamics in the spatial arrangement of species are closely linked to the different demands of species, to their life cycle, mechanisms for reproduction, response to competitive pressures and to the resources available in that system.
Scientific approaches to vegetation dynamics aim to record and explain similarities of vegetation patterns and processes and then to derive characteristics or typical responses and models. The underlying causes of dynamic processes may be found in disturbances of very different kinds. The interpretation of vegetation dynamics is hampered by a confusing multitude of terms, which is caused by insufficient consideration of spatial and temporal scales. The description of vegetation dynamics on a few square centimetres of bare rock will necessarily be different to that within an area of several square kilometres in a forest. For boreal forests a different terminology should probably be used than for tropical rain forests, and taking temporal differences into consideration—hours, days, seasons, years, for example—is probably even more important (Fig. 17.23).

Fig. 17.23. Time scales for dynamic changes in vegetation. (after Miles 1987)
Short-Term and Seasonal Vegetation Dynamics.Plants need to adapt to changing daily and seasonal conditions in their habitat, particularly periodic changes of radiation, day length and temperatures, as well as the amount and distribution of precipitation. The short-term dynamics of plants and vegetation units are considered by the discipline of phenology, the science of seasonal sequences of life cycles of plants and plant communities. Visible changes in the life cycles of plants are recorded, stages of development defined and attempts made to explain these by changes over time, as well as biotic and abiotic factors in the habitat. Of interest are not just typical sequences but also differences in weather conditions from year to year, which may have a considerable effect on the timing of flowering, conditions for germination, the amount and allocation of biomass production and formation of fruit and seeds, as well as the start and duration of individual phe- nological stages.
Phenological characteristics have been known for a long time and are relatively easy to observe. In the daily rhythm, leaves and petals move (heliotropism) in order, for example, to protect against excessive radiation or to capture the highest possible amount of radiation. The production of odorous substances, the secretion of nectar or the supply of pollen by flowers may be coupled with the active phases of animal partners and change during the day. This is mainly regulated by the daily photoperiod.
Different aspects of flowering, harvesting or colouration of leaves are basic observations in phenology. These processes are regulated by photoperiodic responses and light quality. Further, important factors are temperature extremes and temperature integrals or even human intervention. Examples of such controls are the spatial mosaic of vegetation of snow combs, the use of light by spring geophytes in nemoral deciduous forests (Fig. 17.25) or the flowering aspects of meadows and pastures. Plants may require phases of particular low temperatures to germinate (stratification) or for flower induction (vernalisation). Annual, biennial and perennial species behave differently, making the derivation of general rules difficult.
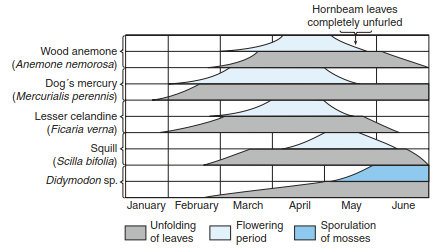
Fig. 17.25 Flowering period of geophytes during spring in a hornbeam (Carpinus betulus) forest. (after Schmidt 1969)
There are also short-term periodic or episodic events that cannot be explained by the usual phenological characteristics but determine the dynamics of these communities decisively. These include regularly occurring floods in river basins, requiring particular adaptations to the extreme water supply and potential restrictions for root respiration (anaerobic conditions) and mechanical disturbance (Fig. 17.26). Another example is the stress caused by avalanches in the Alpine belt.

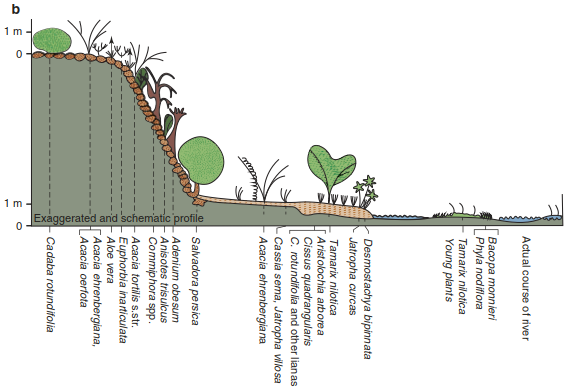
Fig. 17.26. Zonation of plant communities on borders of large wadis in North Yemen (Yemen Arab Republic), determined by different height of water flow. a Wadi Mawr in Tihama Mountains with highly variable water discharge during year (Photo: K. Müller-Hohenstein). b Schematic transect from bed of wadi to lower terraces and to top of wadi bank. Only therophytes such as Bacopa monnieri and Phyla nodiflora are able to colonise the frequently moving gravel banks. In the more strongly consolidated parts of the banks, grasses with particular root systems (rhizomes), such as Desmostachya bipinnata, and very elastic woody species, such as Tamarix nilotica, are able to survive episodic flooding. The resulting changes in vegetation are relatively rapid and aperiodic. (Deil and Müller-Hohenstein 1985)
Phases of activity and dormancy are characteristic of all regions with a seasonal climate. A distinction is made between phases of stronger activity of vegetative and reproductive growth. Occasionally, several vegetative pulses of growth occur, which can be detected as growth rings even in annual species (Schweingruber and Poschlod 2005). In late summer, some tree species rapidly form very distinct new shoots. Seasons may be determined by temperature or from the water supply. In the latter case, seasonal changes in water supply can prove to be the most important regulatory factor for the short-term dynamics of vegetation. In woody species, two types have evolved: Late flowering plants flower only after the development of leaves or after shedding of leaves at the end of the rainy season, while early flowering plants start to flower before the emergence of leaves, i.e. before the rainy season begins. Both types are adapted to the simultaneous occurrence of pollinators or seasonal periods with regular winds.
Because of the usually favourable conditions of temperature and moisture in the perhumid tropics, plants often grow continuously. Nevertheless, irregular bursts of growth may be observed, even with differences among different parts of individual plants. However, definite flowering periods or other phenological phases can hardly be established. Some individuals flower and carry ripe fruits at the same time. It is remarkable that several plant species only flower episodically, sometimes only after several decades.
The timing and duration of specific phenologi- cal phases within a year is very closely correlated with the sequence of weather conditions during the year. These short-term dynamics, occurring year by year, but not necessarily cyclical, are fluctuations. Changes are registered at the level of individual plants. The most important expression of fluctuation is a change in the dominance between co-occurring species. In contrast to suc- cessional dynamics, a return to the previous state is possible. In field studies, it is not easy to distinguish between fluctuation and succession because both overlap and influence each other. The phe- nological spectrum for species of woodland edges for two different annual sequences records the extent of such fluctuations (Fig. 17.24).
In extremely arid regions, where annuals dominate, definite flowering periods no longer occur because of the high variability of precipitation in time and space. But after several rain events a “flowering desert” quickly develops. Many annuals of arid areas may complete their life cycle within a few weeks (Fig. 17.27). The biblical seven fat and seven lean years point to fluctuations and their significance in rain-fed agriculture.

Fig. 17.27. “Flowering desert” (Arabic, acheb) after one of the rare rainfall events in extreme deserts. Such an unpredictable change in vegetation is called a fluctuation; the plant community is composed of only therophytes. This example is from the Arabian Desert (United Arab Emirates) and contains Silene villosa, Senecio glaucus and Plantago amplexicaulis. (Photo: K. Müller-Hohenstein)
Furthermore, fluctuations may occur because of a massive attack by invertebrate herbivores. Not only climatologically but also genetically determined fluctuations are the so- called mast years of woody species with an annual production of a substantial amount of fruit compared to other years. Finally, fluctuations are often the result of human interventions, for example, rotations in grazing management or changing crops in agriculture. The phenological spectrum of species on woodland edges for two different annual sequences records the extent and yearly variability of such fluctuations (Fig. 17.28).
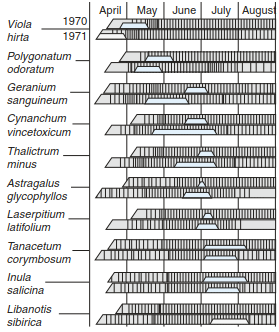
Fig. 17.28. Phenological spectra for species on woodland edges. The small trapezoidal areas show the flowering period. (after Dierschke 1994)
Evaluations of synecological and biocoeno- logical links are only emerging, for example, of the temporal niches of flowering species in plant communities, of the synchronisation of insects and animals that distribute pollen, fruits and seeds. This would not only make pollen calendars available for those with allergies but would also help protect nature, i.e. by providing dates for mowing, for extensive maintenance or as a control for the success of different types of land use.
Date added: 2026-04-26; views: 153;