The characteristics of embryonic development
The embryonic period starts from zygote formation and terminates by birth. The embryonic development includes the following periods: prozygote, zygote, cleavage, gastrulation, tissue and organ formation. The mammalian embryo is called an embryo before formation of the main tissue stems, and is called a fetus after that.
Prozygote period - is the period that precedes zygote development. It was discussed in chapter 4.
Zygote period - it is monocellular stage of new organism development. It is formed as result of sperm and ovum fusion. It was revealed that significant cytoplasm movement in zygotes of Amphibia, Reptilia and Mammalia occurs. Such movements determine regions of further organs and tissue formation (ooplasmatic segregation). The zygote also expresses bilateral symmetry. In the zygote, the protein synthesis starts on a matrix of mRNA made in oogenesis.
Cleavage is a rapid division of the zygote into a larger and larger number of smaller and smaller cells. The pattern of cleavage is greatly influences by presence of yolk. It can be holoblastic (symmetrical and asymmetrical) and meroblastic (discoidal and superficial) (pic 12.1).
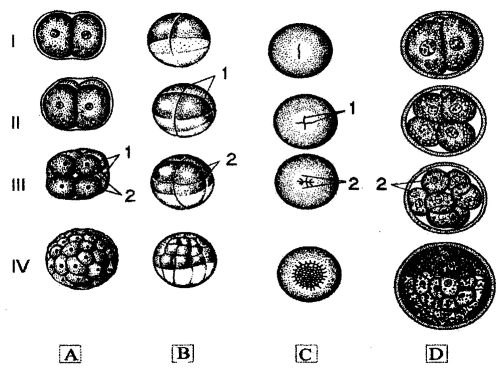
Pic. 12.1. The cleavage of chordates with different ovum type: A - lancelet, В - frog, C - bird, D - mammalian; 1 - two blastomeres, II - four blastomeres, III - three blastomeres, IV - morula; 1 - cleavage grooves, 2 - blastomeres (by N. Yarygin, 1997 with changes)
The symmetrical holoblastic cleavage is in isolecital eggs (in aquatic vertebrates such as lancelets and agnaphants). The cleavage occurs throughout whole the egg. After fertilization, the zygote divides into two cells, which are called blazoners. Then, both cells divide again forming four blastomers. Repeatedly, it increases cell numbers in such line: 2:4:8:16:32, and so on.
The asymmetrical holoblastic cleavage is typical in the telolecital eggs of Amphibia. First two divisions are same as symmetrical division. The cleavage occurs throughout the whole egg too. But yolk-rich cells divide more slowly than those which are poor in yolk. It results in formation of two poles: apical (poor in yolk) and vegetative (rich in yolk). Blastomeres are different in size. Those, which are on apical pole, are smaller than those that are on vegetative pole.
Mammalians and human have little yolk in ovum. They have holoblastic asymmetrical cleavage. Each blastomere has it own rhythm of division. That’s why, the stages of 2, 3,5,7,9 blastomeres can be observed. Some blastomeres are lighter and are placed externally. They give a rise to trophoblast. The cells of the trophoblast can dissolve tissues, perhaps that the embryo can be implanted in the uterine wall. Then, trophoblast cells are separated from embryoblast (darker cells staying intemaly) and make a vesicle. The embryoblast cells are placed on the inner surface of the trophoblasts in shape of disc.
In discoidal meroblastic cleavage, cleavage occurs only in a tiny disc of polar cytoplasm, called blastodisc, which fie astride the large bulk of yolk material. It occurs in the polylecitinal eggs of some mollusks, reptiles, birds and some fish.
Superficial cleavage occurs in centrolecitinal eggs of Arthropoda. The cleavage starts from nucleus cleavage placed centrally in cytoplasm. The nucleuses move outward to regions poor in yolk. The bordering cytoplasm is spited to blastomeres. It results in formation of one layer of blastomeres surrounding yolk material.
In spite of different patterns of cleavage in different organisms, all are terminated by the formation of a blastula. It is one of the signs showing similar origin of fife and parallelism in evolutionary development of structures. At the end of the cleavage, blastomeres are separated by fluid. This fluid localized centrally makes a primary space - blastocoel. Cells of blastula wall are called blastoderm. Starting from blastula, blastomeres are commonly called embryonic cells. All animal species have a blastula stage.
Gastrulation - is the process of two-layer embryo formation. After blastula, all animals start to form layers of embryo. There are four types of gastrulation: invagination, immigration, epibolia and delamination.
Invagination occurs in animals having isolecital eggs. The vegetative pole of blastula invaginates inside. Opposite poles almost touch each other. This decrease volume of blastocoel to minimum and it looks like narrow rime. It results in formation of two-layer embryo. The external layer is called primary ectoderm; the internal layer is called primary entoderm. The invagination form primary intestine (archenteron) or gastrocoel. The opening on one end called blastopore.
Destiny of the blastopore is different. In mollusks, arthropoda, and worms it is transformed into the definitive mouth of the adult organism. Such animals are called Protostomia. In animals having a chorda blastopore it is transformed to an anal canal, whereas the mouth is made on the opposite side as result of complicate processes (invagination of ectoderm and fusion with primary intestine). Such animals are called Deuterostomia.
Immigration was described by I. I. Mechnikov in the embryo of medusa. Some cells of blastoderm migrate to the blastocoel and form a second layer. Both these layers upon being formed surround the gastrocoel.
Epibolia occurs in animals having big eggs rich in yolk (reptilian, birds). Small cells of animal pole are divided quickly than cell of vegetative pole, which is rich in yolk. Cells of animal pole grow over vegetative pole cells becoming external layer. The cells of vegetative pole become internal layer.
Delamination occurs in Cnidarians. It is gastrulation by splitting. During delaminations cells of blastoderm is divided parallel to blastoderm surface. Thus, inner layer underlying ectoderm is made. It was also described by I. I. Mechnicov.
It is important to note that mixed forms of gastrulation may occur too. For example, amphibians have invagination, epibolia and immigration. Only Cnidarians and Sponges terminate their development at a two-layer stage.
All more complicated animals also develop a third layer called the mesoderm. It is situated between the previous two. Mesoderm is of two different kinds: mesenhyme and mesoblast. Mesenchyme is presented by cells immigrated from both ectoderm and endoderm layers. It is spread in the embryo between all the other structures. Mesoblast is formed later. There are two ways for mesoblast formation. One is teloblastic (from Greek “thelos” - end) and second is enterocoelic (from Greek “enteros” - internal, “koiloma” - coel). The first is typical to Protostomia, whereas second is typical to Deuterostomia.
The teloblastic way. It happens when cell groups start to proliferate and migrate inward from both blastopore sides. Then, these groups fill all space between first two layers. Then, cells make secondary coel.
The enterocoelic way. It happens when groups of cells in the form of paired vesicles start to be separated from the primaiy intestine or primary coel. The coel of these vesicles become secondary coelom, which can be segmented. The coelomic vesicles are formed symmetrically from both sides of intestine. The wall of sack looking toward intestine called splanchnopleure, whereas a second one looking toward ectoderm called somatopleure. Thus, the cavities, having very important morphological and functional value, are formed. During formation of the gastrocoel and coelom the volume of the blastocoel significantly decreases. Finally, it transforms to narrow rimes in between intestine wall and coelom. Further, they become spaces of cardiovascular system. Gastrocoel becomes a coel of small intestine. In enterocoelic way, the gastrocoel also gives coelom.
The organs and system formation is a main aim of embryonic period. Embryonic layers contact with each other and this provides connections between different cell groups. Such connections have a great impact on further cell development. They can stimulate each other to develop different signs. Such relations are called embryonic induction. The material of three embryonic layer generate formation of all organs of developing embryo (pic 12.4).
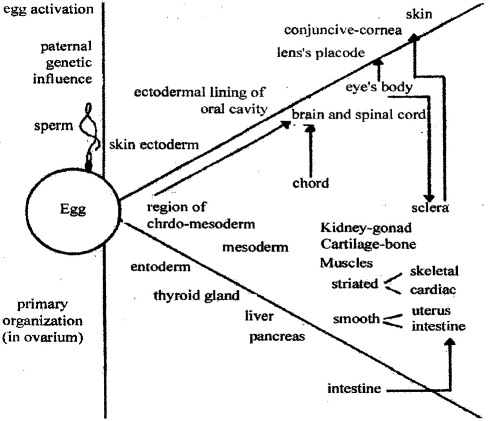
Pic. 12.4. The scheme of vertebrates’ cells differentiation (by B. H. Willier, 1980)
Ectoderm give rise to interguments (external epithelia, skin glands, teethes). A part of ectoderm deeper inside gives rise to the nervous system. Endoderm gives rise to the intestines with digestive glands, and lining of respiratory glands. Mesoderm foiftis all muscular tissues, all types of connective tissues, cartilage, bone, excretory organs, peritoneum, blood, part of ovary and the testis tissue (pic 12.5).
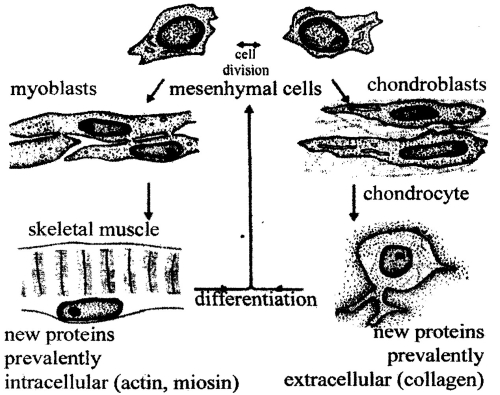
Pic. 12.5. The differentiation of unspecialized mesodermal cells to cells of two types - muscle cells and cartilage cells (by C.H Waddington,1966)
The beginning of organogenesis is called neurulation. Neurulation is the formation of the nervous tube. At the same time secondary intestine and chord are formed. On either side of the developing chord, segmented blocks of tissue form. First, spinal ectoderm induced by chord becomes nervous plate. Then, a layer of ectodermal cells situated above the chord invaginates inward, forming a long groove - neural groove. The edges of this groove then moves toward each other and fuse, creating a long hollow tube, the neural tube, which runs beneath the surface of the embryo’s back. The canal inside the tube called neurocoel (pic 12.8).
Mesoderm separates to dorsal and ventral regions. Dorsal regions are segmented and presented by somites. Ventral part is called side lamina and it connects with somites with help of intermediate mesoderm.
The ventromedial part of somites called sclerotome on being induced by neural tube and chord give rise to vertebrae, bones and cartilages. The intermediate part of somitevcalled myotome give rise to all skeletal muscles. The external dorsolateral pert of somites called dermatome give rise to skin derma. Intermediate mesoderm called nephrotome gives rise to excretory organs and to sexual glands. The primary sex cells separate from other embryonic cells in early development. The mammalians sex cells are supplied by nutrition better than any other cell of the body. Primary sex cells migrate to a place of their definite localization and incorporate in sex glands.
Date added: 2023-01-09; views: 653;