Signalling Networks Involved in Cold Acclimation
Drought, high salinity and freezing stress all pose osmotic challenges for plants. Accordingly, there is considerable overlap in the molecular responses of cells to these different stress conditions. When one is judging by the hundreds to thousands of changes elicited in the A. thaliana transcriptome, however, there are also highly complex responses specific to cold acclimation.
A large number of COR genes belong to the CBF (for C-REPEAT BINDING FACTOR) regulon—that is, they are activated under the control of CBF transcription factors (also termed DRE-Binding proteins (DREB)) (Yamaguchi-Shinozaki and Shinozaki 2006). The promoters of these COR genes, which encode many of the proteins discussed above (i.e. dehydrins, antifreeze proteins, ROS-detoxifying enzymes, fatty acid desaturases, lipid transfer proteins, sugar and proline transporters, osmolyte producing enzymes) share a 5-bp consensus sequence (CCGAC), the so-called Dehydration Responsive Element (DRE), which is known also as Low Temperature Responsive Element (LTRE) or C-Repeat (CRT). Binding of CBFs rapidly activates transcription of COR genes.
The CBF regulon is present in many if not all higher plants (Box 4.5). Its central role in cold acclimation has been demonstrated by overexpression studies. Constitutive strong transcription of CBF genes results in the formation of COR proteins and a permanent cold-acclimated state even in the absence of low-temperature exposure. Such plants can survive transfer to freezing temperatures even without prior acclimation (Jaglo-Ottesen et al. 1998) (Fig. 4.21). CBF overexpression leads, on the other hand, to a strong reduction in the growth rate because CBFs stimulate the production of DELLA proteins, which are central negative regulators of growth (Fig. 4.21b). This is one of many examples for the active dampening of growth as an integral part of stress acclimation responses.
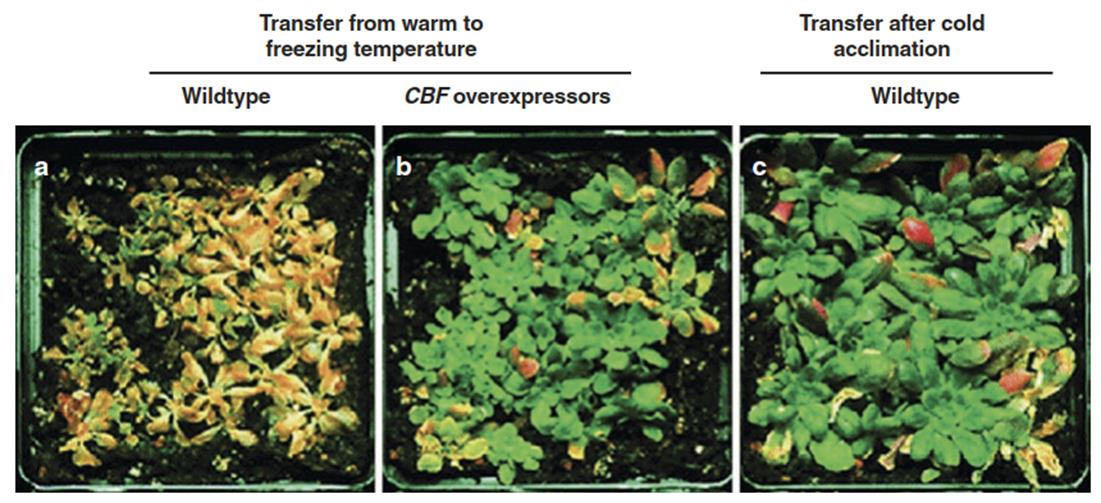
Fig. 4.21. Effect of CBF overexpression on the cold sensitivity of Arabidopsis thaliana. Wild-type plants a require several days of acclimation at low non-freezing temperatures to withstand freezing temperatures c, while plants overexpressing CBF genes survive transfer from warm temperatures directly to sub-zero temperatures b. Note the smaller size of the CBF-overexpressing plants in b. (Modified from Jaglo-Ottesen et al. (1998))
Activation of cold acclimation involves a cascade of regulatory steps that have been unravelled in A thaliana (Fig. 4.22). CBF genes are rapidly up-regulated upon exposure to low non-freezing temperatures. Their expression is controlled by transcription factors that are themselves predominantly controlled at the posttranslational level. The transcription factor ICE1 (Inducer of CBF Expression 1) triggers the transcription of CBF genes by binding to cis elements in the CBF promoters. The stability of ICE1 is controlled by ubiquitination. The ubiquitin E3 ligase HOS1 targets ICE1 for proteasomal degradation, which prevents the expression of CBF genes in the absence of cold. HOS1-antagonistic regulation of ICE1 stability is mediated by SUMO- Ligase-mediated (SIZ1) sumoylation—that is, the addition of SUMO proteins, which protect ICE1 from degradation (Knight and Knight 2012).
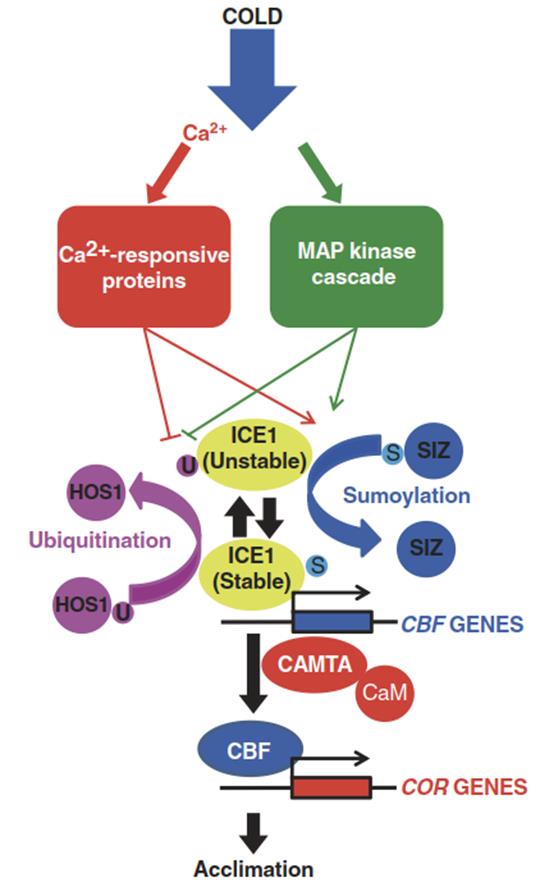
Fig. 4.22. Cold acclimation pathway. Model for signalling leading from cold to the expression of COR genes regulated by CBF transcription factors in Arabidopsis. Transcription factors are represented by solid ellipsoids; other proteins are represented by circles. Red symbolises Ca-dependent processes, green symbolises processes controlled by MAP kinase cascades. S sumoylation, U ubiquitination, CaM calmodulin. (Modified from Knight and Knight (2012))
Events upstream from ICE1 are less well defined. The primary sensor of cold temperature is still unknown. What is clearly established is a rapid increase in cytosolic Ca2+ levels upon transfer of plants to cold conditions, which activates proteins that inhibit ICE1 degradation and activate ICE1 sumoylation. In addition, Ca2+ influences CBF expression via a calmodulin-binding transcription factor (CAMTA). The same regulatory steps leading to enhanced ICE1 activity are triggered by a cold-responsive Ca2+-independent MAP kinase phosphorylation cascade.
Date added: 2025-01-17; views: 542;