Water Use by Plants and Animals
Organisms in their active state do not tolerate desiccation. This is illustrated by the water concentration of tissues: in the active state, the protoplasm of leaves and fleshy fruits contains 0.85-0.90 g H2O g-1FW (fresh weight). The water concentration of wood decreases to about 0.50 g H2O g-1FW due to the high contribution of structural carbohydrates and polymers in the xylem. The lowest values are reached in dormant seeds: 0.05-0.15 g H2O g-1FW.
Land plants must keep the water concentration of their cells close to saturation or be fully saturated in an environment of relatively dry soil or air, and still maintain exchange of CO2 with the atmosphere for photosynthesis. Terrestrial life outside water brings benefits as well as dangers for plants, for the following reasons (Cowan 1977):
- The diffusion coefficient of CO2 in air is about 0.14 x 10-4 m2 s-1 and decreases in water to 0.16 x 10-8 m2 s-1. Thus, CO2 diffuses 10,000 times faster in air than in water (Sestak et al. 1971). In the Lower Devonian, when land plants evolved, the CO2 concentration was significantly higher (about 4000 parts per million (ppm)) than it is today (about 400 ppm), which would have made the atmosphere even more attractive for plants in the past.
- During the evolution of plants, no membrane has been “invented” that is permeable to CO2 but remains impermeable to H2O vapour. Even in the future, there will not be a type of “GoreTex” for CO2, because the molecular weight of CO2 is larger than that of H2O (44 versus 18). Of course, it was the availability of CO2 in the atmosphere as a resource that made plants adapt from life in water (algae) to life on land. However, in order to use this carbon supply, mechanisms had to be developed to regulate the cellular water relations of land plants.
- water relations are more important for the gas exchange of land plants than for animals because of the chemical composition of air. Photosynthesis creates a CO2 gradient of about 100 ppm between the atmosphere and the mesophyll. At the same time, there is a gradient of water vapour of about 12,000 ppm between the water-saturated mesophyll walls and the ambient air. CO2 diffuses 1.6 times more slowly than H2O vapour, as the diffusion rate is related to the square root of the molecular weights

Thus, during uptake of 1 mole of CO2, the plant loses about 200 moles of H2O 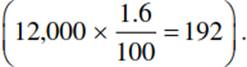 Therefore, water use (here, water loss) by plants is very high relative to the photosynthetic gain.
Therefore, water use (here, water loss) by plants is very high relative to the photosynthetic gain.
Water use by animals differs from its use by plants. For mammals (warm-blooded animals with a body temperature of 37 °C), with 210.000 ppm O2 in the atmosphere and 160.000 ppm O2 in the breath, the O2 gradient is about 50,000 ppm. The air that is breathed out is water saturated. Therefore, the water vapour gradient between the lung and the atmosphere is also about 50,000 ppm (at 20 °C, 50% rel. air humidity). Thus, a warm-blooded animal loses only about 1 mole of H2O per mole of O2 taken up. For cold-blooded animals the water loss related to O2 uptake is even lower (about 0.2)—that is, water use related to O2 gain is very low in animals. In addition, animals can move to a water source or protect themselves from adverse conditions, giving animals another advantage over plants under dry conditions.
As the loss of water from plants is so large, it is usually not cost effective for them to have water storage. During the course of a day, a sunflower leaf loses about ten times its own weight as water vapour. A 25 m high spruce loses about 100 – 1000 L of water per day, which is more than there is in the stem. The available water stored in the stem and the crown of spruce is sufficient to maintain transpiration for only about 2 h in the humid morning hours (Schulze et al. 1985). This means that the costs of providing water from storage sufficient to safeguard the supply over days or months would be unreasonable: plants would have to construct enormous water stores.
The aforementioned example of the daily water loss by a spruce tree shows that a 10-day drought period without water uptake from the soil would require a storage volume of up to 10,000 L to support normal transpiration. There are some species of cacti, euphorbias and Mesembryanthemaceae that live transiently on water stored in cell vacuoles (e.g. Oppophyllum spp. or Prenia spp.), but their biomass production is then very low. Even the baobab tree (Adansonia digitata) is hardly able to use the water stored in its stem (Schulze et al. 1998b). The wooden structure of baobab and other “bottle trees” is rigid and cannot shrink sufficiently to maintain transpiration. The function of the thickened stems of baobab trees is to store carbohydrates and amino acid reserves in sufficiently hydrated cells.
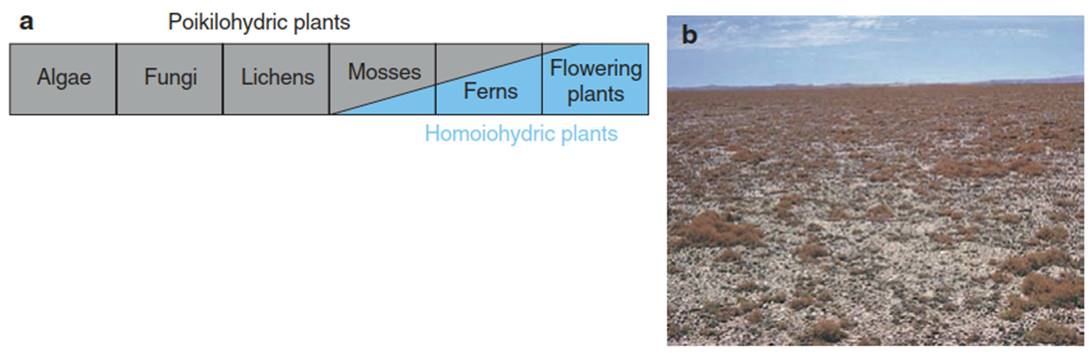
Fig. 10.1. Desiccation-tolerant and -intolerant plants. (homoiohydric) organisms. b Lichen growth dominated a Schematic presentation of the occurrence of desiccation- by Teloschistes, with almost no higher plants in the coastal tolerant (poikilohydric) and desiccation-intolerant region of the fog desert of Namibia
During plant evolution, two strategies of water use have developed (Fig. 10.1a):
1. Desiccation-tolerant, poikilohydric plants operate as a physical system of variable water content, which absorbs and loses water, depending on the humidity in the air. When exposed to moisture from rain, dew or high humidity, these plants become fully active. They dry out with decreasing air humidity and become dormant during the time of desiccation. In this life cycle, there is an optimum water status. If the organism is too wet, diffusion of CO2 from the atmosphere to the photosyntheti- cally active cells is restricted and the rate of photosynthesis decreases with high water content (see Fig. 12.13). Thus, metabolism is restricted to the period when the tissue is wet, but not too wet, and diffusion of gases is possible. The most important representatives of this plant type of water use are algae, lichens and mosses (Fig. 10.1).
Some mosses and lichens are able to obtain water from the ground by rhizoids and fungal hyphae via capillary forces but not via xylem vessels. Among flowering plants, there are also a few species that are desiccation tolerant—for example, Myrothamnus flabelliformis from the Namib Desert; Borya spp., native to Australia; and Craterostigma plantagineum in Africa, as described by Ziegler and Vieweg (1970) and Gaff (1971). These angiosperms, however, differ from non-vascular plants in that they cannot be activated by dew and high humidity as they have a cuticle, which restricts water uptake via the shoot (see also Burkhardt (2010)). These species are activated by water uptake through roots only.
2. Desiccation-intolerant, homoiohydric plants are able to maintain a high and almost constant tissue water content that is independent of the conditions in the surrounding environment. There is a partitioning of labour between organs (Fig. 10.2): roots are specialised in uptake of water, the stem transports water and the green tissue (which may be leaves, phyllodes or phyl- loclades) assimilates CO2 at the cost of evaporation. Homoiohydric plants have large vacuoles in their cells, which function, within a certain range, as short-term buffers for the cellular water status and thus stabilise the cell and plant water balance.
The leaf surface is covered with a cuticle composed of a lipophilic polymer, which is impermeable to CO2 and H2O but permeable to O2. The leaf is connected to the free atmosphere via stomata, of which the aperture can be regulated. However, one developmental stage of intensive dehydration of cells also remains in these homoiohydric plants: the seed. In environments with high air humidity and low rainfall (e.g. the fog desert of the Namib, where there is no precipitation except for dew), higher plants are inferior— with respect to cover and growth—to lower plants (Fig. 10.1b).
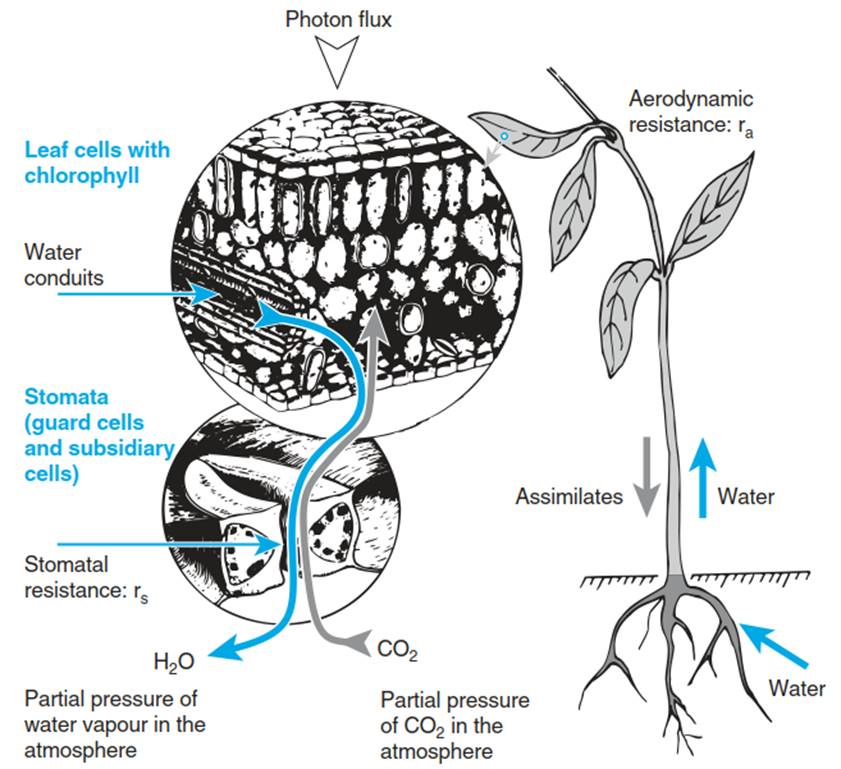
Fig. 10.2. Flow of water and assimilates in vascular plants. These are connected via the stem, which conducts the water and assimilate flows. In the stem, water and assimilates are transported by mass flow; in the leaf, there is a phase transition from liquid water to water vapour. At the same time, CO2 is assimilated into soluble organic substances. Diffusion between the inside of the leaf and the atmosphere is controlled by the stomata
In this chapter, our focus is on homoiohydric plants, as they form the largest fraction of the terrestrial flora. While Chap. 6 presents the molecular basis for the responses of plants to water stress, this chapter will focus on the biophysical links between plants and the environment.
Date added: 2025-02-05; views: 639;