Water Transport from the Soil to the Plant
Water Uptake. Precipitation seeps through the soil profile and may reach groundwater. The amount and rate of water movement in the soil depends on the soil type, the pore size and the water saturation. The field capacity describes the content of water, which is retained against gravity—that is, it is the amount of water that does not drip out of a flower pot after watering (field capacity is lower for sand than for loam). However, this water is only partially available for plants, because a proportion is bound as hydration water or by capillary forces— the matrix potential (ψm) of the soil. In pores of 5 nm (e.g. pores in secondary cell walls of higher plants), the matrix potential reaches 3.0 MPa. It decreases in pores with a radius of 500 pm to only 30 Pa (Schachtschabel et al. 1998). Threshold values have been defined for the characterisation of soils on the basis of the relation between the water potential and the soil water content (Fig. 10.6a) (Or et al. 2012).
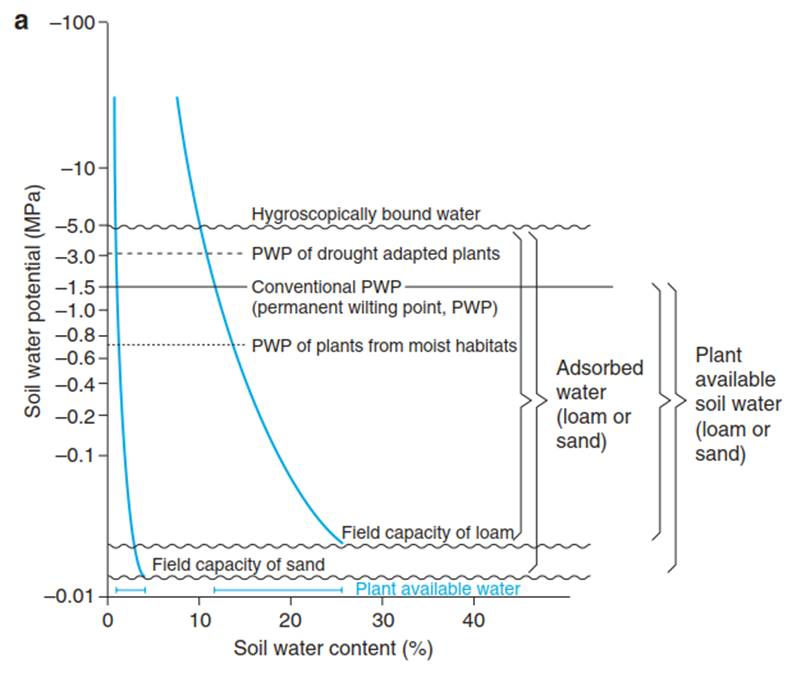
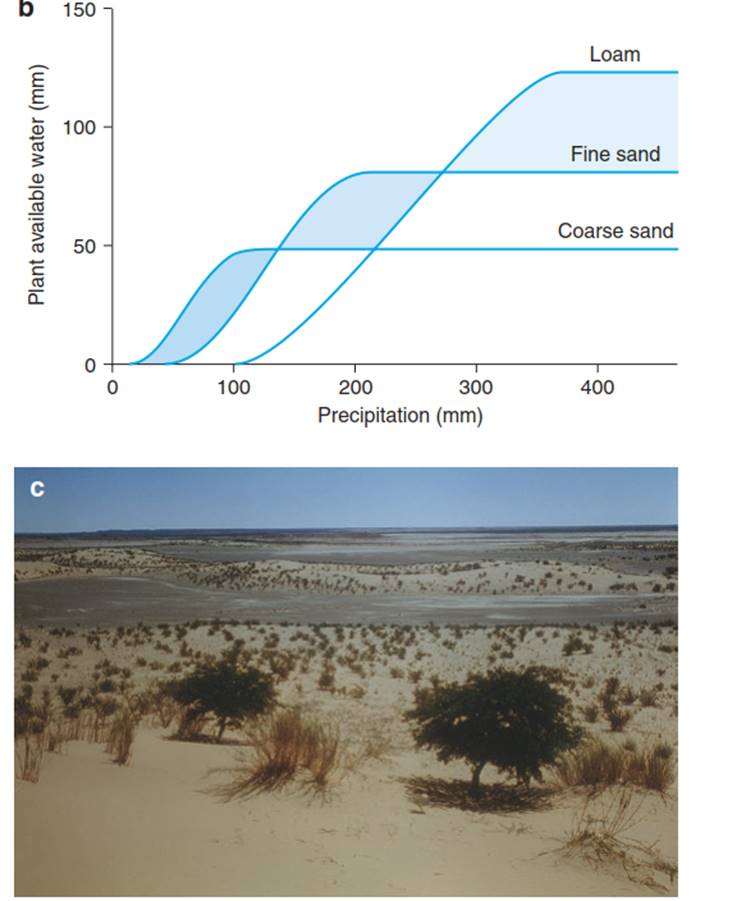
Fig. 10.6. Changes in water status and plant-available water in sandy and loamy soils. a Depending on the texture, the water potential at field capacity is -0.05 MPa (in sand) and -0.015 MPa (in loam). Convention sets the permanent wilting point (PWP) of agricultural systems at -1.5 MPa. Plants that live in wet or dry conditions may have wilting points at -0.7 MPa and -3 MPa, respectively, but the additional available water resulting from a shift in the wilting point is small. The limit of hygroscopically bound water is set at -5 MPa.
The difference between field capacity and hygroscopically bound water is the exchangeable water, only a proportion of which is available for the plant (from Larcher (2003)). b Change in the amount of water available to a plant, depending on the soil texture. In arid areas, sandy soils contain more water that is available to a plant than in a loamy soil, as the water is not as strongly held by capillarity in sand as it is in loam. In contrast, in areas of high rainfall, the amount of water that can be stored by a sandy soil is less than that in a loamy soil (Modified from Walter (1960)). c Distribution of vegetation in the Namib Desert (Asab, Namibia) with Acacia karroo, Aristida sabulicola and Acacia ciliata growing on the sand dunes (with high water availability and low salt) and vegetation-free areas on the plain, where loam and clay soils have low water availability and higher salt concentrations
Loam and clay soils differ not only in the amount of available water (for loam it is almost ten times higher than for sandy soils), but also in the amount of water that is not available for plants in dry soils. This has consequences for the plant availability of water with decreasing precipitation (Fig. 10.6b). In areas with high precipitation, sandy soils are drier habitats than loamy or clay soils because sandy soils retain less water, owing to the larger pore sizes. At low precipitation the limit at which water can no longer be taken up by the plant is reached earlier in clay because the capillary and matrix forces are reached sooner than in sandy soils. In arid regions, sandy soils are therefore “moister” habitats for plants than clay soils. This is modified in nature by the frequency of precipitation (Fig. 10.6c).
Water transport from the soil to the root is determined not only by the water potential gradient but also by the hydraulic conductivity, which depends on the soil water content and soil texture. Roots may grow towards the water or “wait” until water flows from the soil to the root. In most plants, water transport to the roots is additionally facilitated by fungal hyphae forming the so-called mycorrhizae (Marjanovic and Nehls 2008), in which water is mainly transported towards the root by capillary forces. The availability of water in the soil determines the surface area of roots needed to provide leaves with a certain amount of water.
Water transport in soils is different in saturated soils (water content above field capacity) and unsaturated soils (water content below field capacity; water moves as vapour). In saturated soils, water flow follows Darcy’s law (Darcy 1856). Darcy studied the water pressure in the wells of the city of Dijon, France. In this case the flux, or Darcy velocity (v, in volume per time and area, measured in cubic metres per hour per square metre or in metres per second), is proportional to the hydraulic gradient (dh/dx: change in height per change in the length of the flow path, which corresponds to the potential gradient) and the saturated soil water conductivity (ks), which depends on soil texture, where soil texture describes the grain size of soil particles (stone, sand, silt, clay):
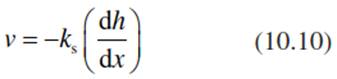
In stony soils к is >10-3 m s-1, in sand it is >10-5 m s-1, in silt it is 10-7 m s-1 and in clay it is 10-9 m s-1. In unsaturated soils, the rate of flux is much lower than that in saturated soils. It depends on the unsaturated soil water conductivity, k0, which decreases with the soil water content, θ, and is determined by the potential gradient, ∆ψ, over the distance, x.
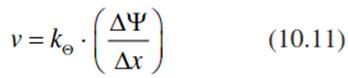
This equation is analogous to Eq. 10.10, but the water potential is the driving force, which in turn is related to the relative humidity in soil pores. The values for kθ in silt soils range between 10-13 near saturation and 10-17 cm s-1 close to the wilting point. Soils are not homogeneous but structured in horizons and differentiated within the horizons in more or less dense aggregates. Thus, the hydraulic conductivity of soils varies over a very short distance with texture, aggregation and soil water content (Horn 1994). Thus, even within a single soil profile, water availability is highly variable, which results in high variability in the root architecture of plants. In dicots, the seminal root is long lived and, as a tap root, explores deeper soil horizons with an extensive root system. In horizons where nutrients and water are available, a dense adventitious root system is formed. In contrast, in grasses the seminal root is short lived. A dense adventitious root system develops near the soil surface (Fig. 10.7). There is an interaction between water and nutrient availability on the one hand and the hydraulic structure on the other (Ewers et al. 2000).
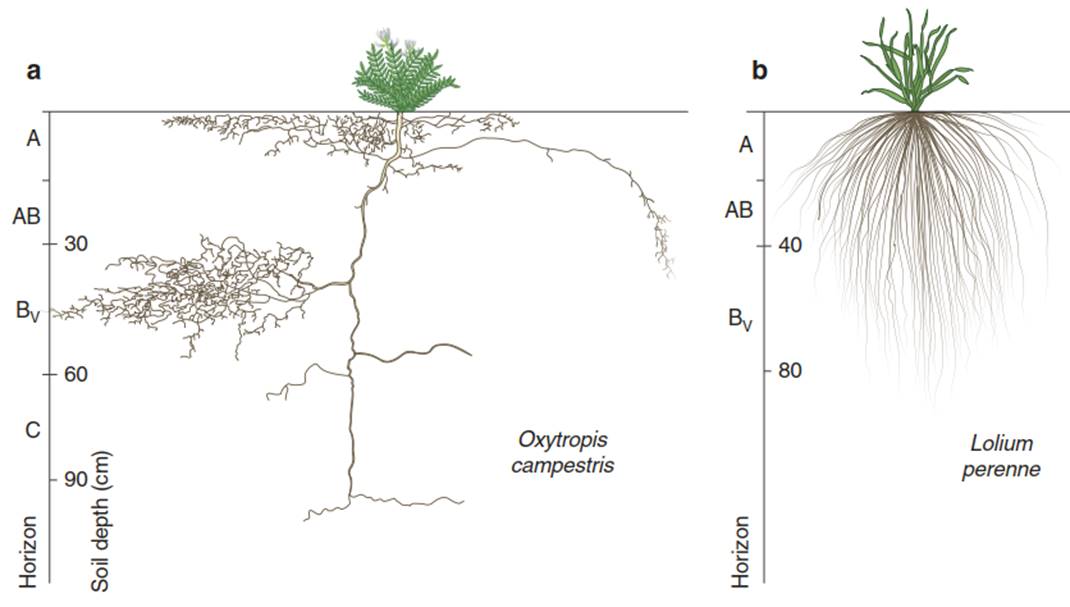
Fig. 10.7. Root architecture of dicotyledonous and monocotyledonous plants. Root distribution of a the dicotyledonous Oxytropis campestris and b the grass Lolium perenne. (redrawn from Kutschera and Lichtenegger (1992))
The soil layer from which plants gain their transpiration water may be determined with stable isotopes of hydrogen, D = deuterium, and oxygen, 18O. The stable isotope values for hydrogen and oxygen (δD and δ18O) increase with the temperature of precipitation (Dawson 1993)— that is, winter rains have a lower stable isotope value than summer rains. In addition, isotopes enrich at the soil surface, as water molecules containing heavier isotopes evaporate more slowly than molecules with lighter isotopes. Measuring the isotope ratios in (D/H or 18O/16O) xylem water and in water along a soil profile provides information on the horizons from which species may take up their water.
Establishing the origin of transpired water (water uptake) with reference to the soil depth is most clear in regions with seasonal rainfall at different temperatures. An example is given in Fig. 10.8a for the Colorado Desert in the southwestern USA (Ehleringer 1993, 1995). Summer and winter rains show very different 8D values, and groundwater is even more depleted in deuterium than winter rain because of isotope fractionation during seepage of water to a greater depth. Various plant species use water sources from different depths. In this subtropical “warm” desert (Fig. 10.8b), annuals and succulents use summer rains, which reach the Colorado Desert as sporadic subtropical fronts (note: in Mediterranean winter-rain regions, annuals use mainly winter rains). In contrast to the summer annuals, deeper- rooting perennials (usually, evergreen herbaceous plants and shrubs) utilise the water of winter rains or groundwater.
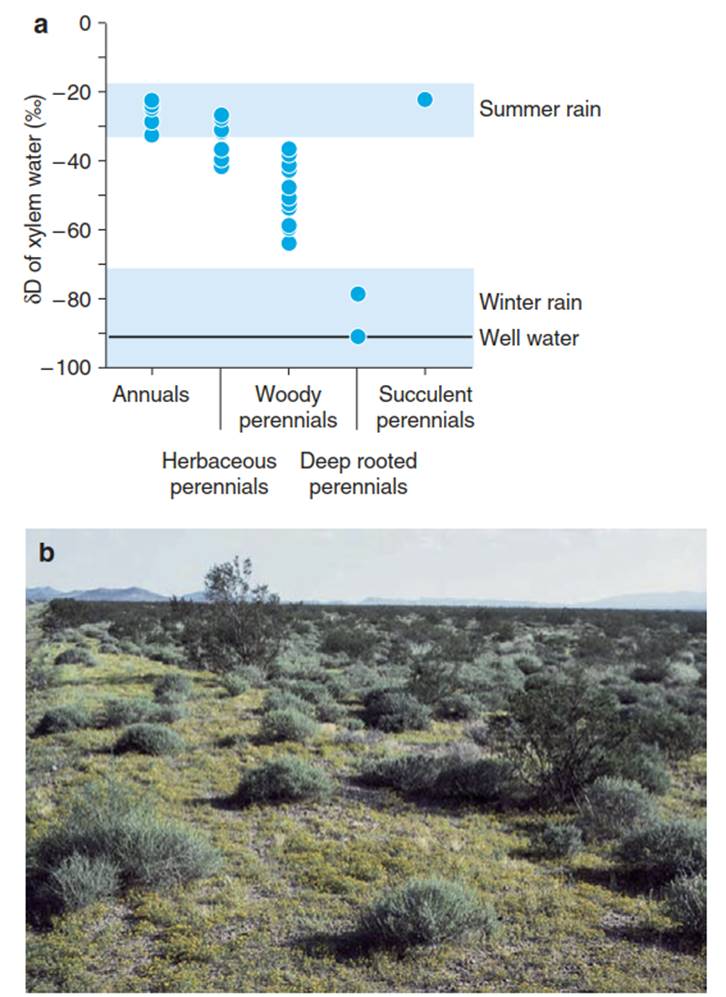
Fig. 10.8. Stable isotope values of hydrogen for plant types and groundwater in the Sonoran Desert. a Hydrogen isotope values in water from the xylem of different types of plants growing in the Sonoran Desert (blue circles), compared with the hydrogen isotope values in summer and winter rainfall (blue bands), as well as in groundwater (black line) (after Ehleringer (1993)). The 5D value is calculated from the D/H ratio of the sample in comparison with a standard [(D/H)sample/(D/H)standart - 1], where water from deep oceans is used as the standard (V-SMOW = Vienna standard mean ocean water).
In the case of the Sonoran Desert, annuals use the summer rain water, which has a high δD value depending on the temperature, almost exclusively. In contrast, deep-rooted perennials almost exclusively use water from winter rainfall, which has a low δD value due to the lower temperatures. The δD value of water from the xylem of plants shows from which soil level they obtain the water. b Vegetation in the Sonoran Desert close to Oatmans, South Nevada, with summer annual plants Pectis papposa (Asteridae, C4 plants); perennial woody plants, Ambrosia dumosa (Asteridae, C3 plants); and perennial deep-rooted Larrea tridentata (Zygophyllaceae). (Photo: E.-D. Schulze)
Date added: 2025-02-05; views: 560;