Plant Pollination: Vectors, Floral Traits, and Specialization
Wind and animals are the most important vectors for pollen. Pollination by wind can occur over greater distances, but the success declines remarkably with distance. The distance of pollen transported by animals is usually much more restricted, but pollen carried by animals often has higher chances of ending up on the stigma of a conspecific. This is because animals do not move randomly within an area but rather direct their movements to locate food resources thereby increasing the probability that pollen will be deposited on flowers.
The key element in pollination is thus that pollen needs to be moved from one flower to the next conspecific one for successful pollination. To enable that movement, pollen must stick to animals, for example, the hairs of insects or feathers of hummingbirds, and be brushed off on the stigma of the next flower. Flowers typically offer rewards to entice animals to visit them. The most common types of rewards are nectar and pollen; less common rewards include resins, fragrances that insects collect in specific grooves and heat. For example, male orchid bees (Euglossini) collect fragrances from flowers and other resources and store them in specialised tibia pockets to use them later in courtship (Eltz et al. 2008). In some cases, insects visit flowers not for their nutritional rewards but because they have a high likelihood of finding mating partners on flowers.
Not all animals visiting flowers to collect rewards are legitimate pollinators; some leave flowers without having transferred pollen or taken it up. To complicate the issue further, species such as bumblebees or hummingbirds that may be legitimate pollinators in one plant species may be nectar thieves in another. To study pollination systems, it is thus essential to verify the effect of animals on pollination.
Flowers that exceed the mean ambient temperature can offer considerable metabolic rewards in terms of heat to ectothermic insects. Self-heating flowers evolved independently in 6 families, with an estimated number of 900 species alone being pollinated by scarab beetles of the genus Cyclo- cephala. Seymour et al. (2003) showed that beetles resting inside 3-4 °C warmer flowers had two- to five-fold lower metabolic rates than those outside the flowers. Thermogenesis is associated with elevated respiratory rates, which is why CO2 emission can be a reliable indicator of heat production. Because insects spend a considerable amount of time in warm flowers, there is a high probability of successful pollen transfer. Plants can also increase the probability of pollen transfer by trapping insects until pollen has been transferred.
Because floral rewards can be unpredictable and are often not immediately visible to approaching floral visitors, many pollinators learn quickly to associate floral phenotypes with the expected average rewards. Learning of variable phenotype-reward associations can lead to an important behaviour for plant fitness, floral constancy. Floral constancy describes the behaviour of preferentially visiting one or a few rewarding floral phenotypes in subsequent visits. Flower constancy is an important behaviour that leads to a high probability of the arrival of conspecific pollen. Hence, flowers should be under selection to be memorisable for animals, a selection that is not necessarily found in fruits since successful seed dispersal does not require animals to visit conspecific plants.
Because plants have a high interest in not losing pollen on flowers of different species, interactions with pollinators are on average more specialised than are those with seed dispersers. The higher specialisation is explicable because floral phenotypes represent important filters that structure the interactions between flowers and floral visitors. Perhaps the most important filter is floral morphology, which can severely restrict the ability of animals to reach the nutritional rewards of the flower. In addition, floral morphology obviously influences not only the likelihood that pollen will be transmitted by a floral visitor but also the sequence of events. Presenting pollen and stigmata that receive pollen so that pollinators touch them before they can exploit nutritional rewards (referred to as approach herkogamy) can be adaptive because pollinators provide their pollination service before plants deliver payment in nectar.
This situation contrasts with that of seed dispersal, where plants have no control over the movements of animal vectors once they leave the fruiting plant. Here, plants necessarily pay in advance—in terms of the energy provided by fleshy fruits—prior to the seed dispersal event. Third, flowers of a given species are under greater selective pressure to be recognisable and visited repeatedly than the fruits of the same species. Consequently, phenotypes that are important in species recognition such as colours and scent are more diverse in flowers than they are in fruits. For example, the diversity of floral colours is twice that of fruit colours, and flowers sport colour patterns more often than fruits, which creates further diversity in visual floral displays (Stournaras et al. 2013).
Even though pollination systems are more specialised than seed dispersal systems, there is strong variation in the degree of specialisation. On one end of the generalisation-specialisation continuum, only a single species pollinates flowers, whereas other plants are pollinated by more than 50 different species. Figs are often quoted as a remarkable example of a particularly specialised pollination system. Bertin (1989) described the cycle for Ficus sycomorus (Fig. 19.28). The syconium, the large, almost closed inflorescence of this fig species, has a small distal opening, the ostiulum. Females of one group of wasps from the family Agaonides are attracted by the smell of the flowers and carry pollen through this opening into the inner space (in one direction only).
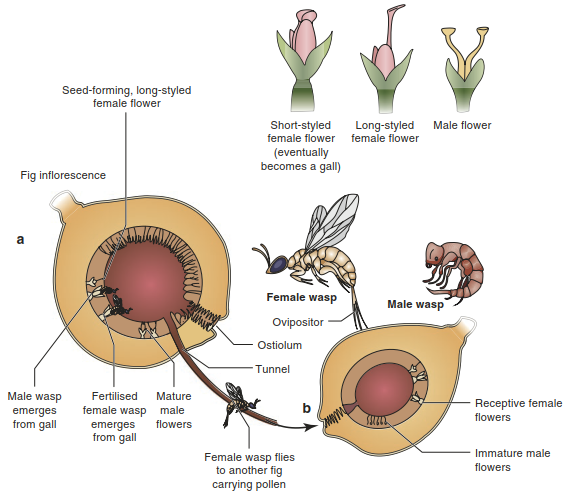
Fig. 19.28. Ficus sycomorus and its wasp pollinators
They distribute pollen to the numerous individual female flowers in the inner space. In some of them, particularly those with shorter pistils, they lay eggs before they die. At the same time, individual figs develop and the larvae of the fig wasp feed on the gall-like growing fig tissue. First male wasps hatch and find galls with females and fertilise them. The males die or dig tunnels and reach the outside. The females pass through these tunnels to the outside, taking up pollen near the tunnel. This is a very specialised pollination system with coevolutionary, morphological and physiological adaptations of wasps and figs.
There appears to be a latitudinal gradient, with the pollination system of tropical species being on average more specialised than that of temperate species. Floral morphology is key to understanding the degree of specialisation. Open, radial symmetric flowers (actinomorphic) are less efficient at directing pollinator movement within flowers to the stigmata and thus often exhibit a “generalised pollination system”. In contrast, closed zygomorphic flowers with bilateral symmetry restrict the directionality of approach and movement within the flowers, leading to more efficient pollen transfer. Radial symmetry is the ancestral form, with zygomorphic flowers having evolved independently several times. The length of flower tubes is a further morphological trait that can strongly influence which animals are able to enjoy the floral rewards. The longer the tubes, the more restricted and, hence, specialised the pollination system will be.
The most important function of floral morphology is to ensure effective pollination. As discussed earlier, this can partly be achieved by screening out nectar thieves or unreliable pollinators. Unreliable pollinatores are those visiting a species only irregularly and thereby increasing the risk of losing pollen. However, plants can also increase pollination effectiveness by guiding pollinators to the reproductive organs. It is here where flower morphology is particularly important. For example, Muchhala (2007) showed that bats and hummingbirds selected for different corolla widths. In an experiment, he showed that wide corollas guided bat snouts better, whereas narrow corollas guided hummingbirds’ bills better. A poor fit between floral morphology and pollinator morphology resulted in variable entry angles and decreased pollen transfer. Interestingly, generalisation in plant traits was never favoured in this experiment because intermediate corollas always had lower pollen transfer rates.
Nectar guides are another morphological feature to help pollinators orient on flowers. Nectar guides can be morphological, for example, elevated rims on the surface of the petals, or visual, such as coloured marks or UV stripes radiating from the floral rewards (Fig. 19.29). There is also possibly an olfactory correspondent of nectar guides because flower parts of the tall buttercup (Ranunculus acris) differed in odour profiles (Bergstrom et al. 1995). Most remarkably, pollen odour differed strongly from all other plant parts, and emission from the petals differed among apical and basal petal regions, paralleling optical nectar guides.

Fig. 19.29. Plants guide pollinators to their reproductive organs. Nectar guides are morphological or visual traits such as rims that radiate from the centre of a flower and help pollinators to orient on it and find the nutritional rewards
Earlier, we stated that animals typically visit flowers to collect nutritional rewards. Yet flower visitors need to be tolerant to variable nutritional returns. Even in highly rewarding species that offer abundant nectar or pollen resources, the flower may have been depleted by recent visits of competing pollinators. Pollinators thus need to be tolerant to variable nutritional returns linked to a given floral trait. This tolerance is important in evolutionary terms. It facilitates, for example, the repeated evolution of rewardless flowers precisely because tolerance leads to weaker selection against rewardlessness. Although flower species from 32 families are permanently rewardless (Renner 2005), detailed data are available only for orchids, which are the prime example of deceptive flowers, with roughly a third of all species not offering any rewards to pollinators. To achieve pollination, the most commonly employed strategies of rewardless orchids are food and sexual deception.
Food-deceptive species may mimic rewarding species or rely on perceptual exploitation of innate biases in the sensory system of pollinators. For example, a widespread innate bias of pollinators is that they preferentially visit large floral displays. Large floral displays are more easily detected from a distance, but the preference for large displays persists even in lab conditions where distances are typically small. Sexually deceptive orchids are probably the prime example of mimicry in plants. They mimic the scent of female pollinators and entice males to attempt to copulate with the flower, which also provides some visual resemblance to female pollinators. According to Jersakova et al. (2006), food deception is six times as common as the more specialised sexual deception in orchids. An important consequence of rewardlessness is an increased outcrossing rate as pollinators leave unrewarding plants more quickly than rewarding plants. As such, they may visit fewer flowers of the same individual, which means the pollen of the flowers they did visit will be disproportionally deposited on the flowers of different individuals.
Although pollinators need to be tolerant to variable nutritional returns, there can also be selection for more reliable signals. For example, flowers of at least 77 families change colour once they are pollinated (Weiss 1995). These unrewarding flowers still contribute to attracting pollinators from a distance to as yet unpollinated flowers of the same plant. At the same time, they also increase the foraging efficiency of pollinators by directing them to rewarding flowers. The repeated evolution of the trait of colour change is thus thought to originate from a coevolutionary process between plants and their pollinators that increases the efficiency of pollen transfer.
Flowers and pollination are important in premating isolation and speciation in plants. In particular, pollinators can drive speciation in plants if they restrict their visit to specific phenotypes. This is termed ethological isolation, and it can constitute an important premating isolating barrier. The proximate basis for ethological isolation is the distinct sensory world of pollinators. Ethological isolation is typically studied in hybrid zones, for example, that of Ipomopsis aggregata and I. tenuituba. In this hybrid zone, ethologi- cal isolation mediated by difference in flower colour was more important than mechanical isolation owing to distinct floral morphologies. I. aggregata is characterised by red flowers with wide corolla tubes and a high nectar production, whereas I. tenuituba has white to pink flowers with long and narrow corolla tubes and low nectar production. Painting flowers of both species red increased the percentage of conspecific seeds (Campbell and Aldridge 2006), demonstrating the importance of plant-animal communication in directing gene flow.
Although the concept of diversifying selection exerted by pollinators upon floral traits is intuitively plausible, and although this process has likely played an important role in the diversification of angiosperms, it remains to some extent controversial. While interspecific comparisons consistently suggest that animal-pollinated clades are more species-rich than clades with other pollination systems, it has been difficult to exclude the possibility that this pattern is caused by animal-mediated pollination rather than other selective agents not controlled for in analyses.
Odours are likely to be the primary communication channel of higher plants because fragrances were nearly universally employed among the early angiosperms. However, olfactory communication by plants is not as well known as visual communication, partly reflecting our own bias as visual animals, partly reflecting the more complex nature of olfactory signals. For example, more than 1700 substances are known from floral scent (Knudsen et al. 2006). However, many of these substances are not known to function in attracting pollinators. For example, only 9 of more than 60 volatiles in sacred datura (Datura wrightii) elicit a behavioural response in its pollinator, the moth Manduca sexta (Riffel et al. 2009). The complexity of odour blends means that it is often not a single substance that attracts animals but a mix of various compounds. In a few pollination systems, odours constitute the reward, such as in Euglossini bees that collect perfumes from flowers (Eltz et al. 2008).
It should be emphasised that pollinators use both floral odour and colour to identify and discriminate among flowers. Visual and olfactory stimuli can have additive effects on pollinators. The additive salience of visual and olfactory stimuli was a good predictor of bumblebees’ initial behaviour and learning performance of floral stimuli (Katzenberger et al. 2013). Importantly, this study showed that floral displays consisting of olfactory and visual stimuli with low individual salience were more effective than the summed predicted compound salience, pointing to a multiplicative effect of the combination of olfactory and visual signals. In other words, the simultaneous integration of olfactory and visual stimuli disproportionately increased perceivers’ discrimination abilities of less efficient stimuli. This effect was not identical for vision and scent in bumblebees: Multimodal floral signals increased learning relative to only olfactory cues, but not relative to only visual cues. Hence, this and other studies document that pollinators select not a single trait but a combination of different traits, which has been termed a pollination syndrome.
Date added: 2026-04-26; views: 165;