Regulation of Avoidance and Tolerance Strategies
The metabolic and morphological plasticity of flooding-tolerant plants enables a wide range of survival strategies, of which escape and quiescence can be regarded as the extreme poles. Molecular dissection of these contrasting strategies in rice has revealed that they are regulated by very similar factors. This provides direct insights into how, during evolution, successful adaptation to a variety of habitats can arise within a species or a genus.
Several low-yielding rice landraces cultivated in flood-prone areas carry loci that are responsible for the rapid shoot elongation trait (SNORKEL1 and SNORKEL2 (SK1 and SK2) (Hattori et al. 2009)) or the energy-saving mode. Encoded by these loci are group VII ethylene response factors—that is, members of the family that in A. thaliana has been shown to be regulated by the oxygen-sensing pathway (Fig. 5.13).
Thus, the integration of the ethylene signal and the O2 sensing is utilised to trigger very different responses to oxygen deprivation through variation in the targets of ethylene and low-oxygen signalling. The underlying circuitry and its recruitment for contrasting strategies exemplify the central role of differential growth control for the adaptation to stress conditions.
Shoot growth is dependent on the plant growth hormone gibberellic acid (GA). One of its classic activities is stimulation of internode elongation. Accordingly, the SNORKEL genes stimulate the GA response (Fig. 5.14). In addition, ethylene triggers the decrease in the endogenous abscisic acid (ABA) level and thus in turn increases the effectiveness of GA. Part of the GA response is the induction of expan- sins—small proteins that are involved in the loosening of hydrogen bonds between cellulose and hemicelluloses in the cell wall.
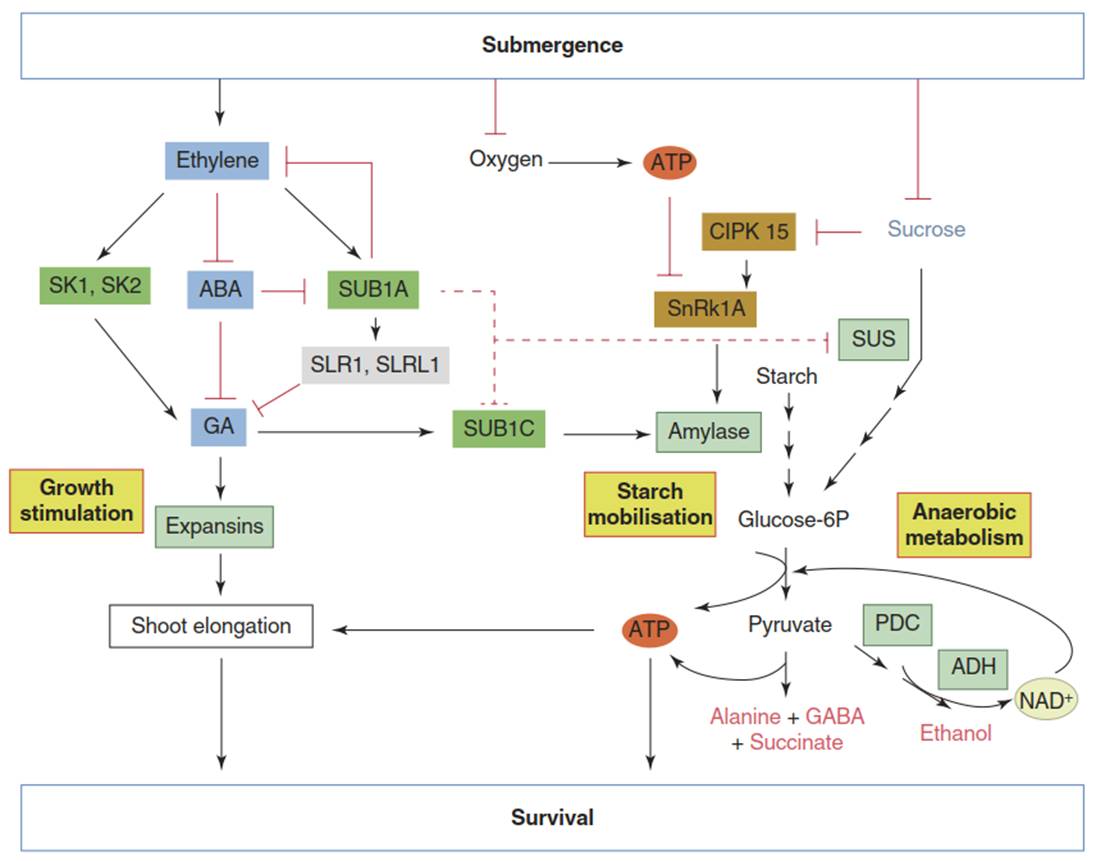
Fig. 5.14. Contrasting submergence survival strategies are mediated by ethylene response factors in rice accessions. Upon submergence of aerial organs, ethylene concentrations rise, adenosine triphosphate (ATP) becomes depleted and sucrose is rapidly consumed. These events trigger contrasting strategies in different rice genotypes: either promotion of elongation growth in deepwater rice (escape) or repression of growth in accessions tolerant of deep submergence (quiescence). The escape strategy is controlled by the ethylene response factors SNORKEL1 (SK1) and SNORKEL2 (SK2). They stimulate gibberellic acid (GA)-dependent elongation of internodes. Among the activated proteins are expansins.
GA also induces another ethylene response factor, SUB1C, which activates starch mobilisation and the provision of ATP through anaerobic metabolism (Fig. 5.11; ADH alcohol dehydrogenase, PDC pyruvate decarboxylase, SUS sucrose synthase). Depletion of ATP and sucrose pools trigger these metabolic processes through kinases such as CIPK15 and SnRK1A. By contrast, in rice genotypes carrying the SUB1A gene, ethylene leads to an increased accumulation of the GA response inhibitors SLENDER RICE 1 (SLR1) and SLENDER RICE-LIKE 1 (SLRL1). Furthermore, the consumption of sucrose and starch, as well as the synthesis of ethylene, are inhibited. (Modified from Bailey- Serres and Voesenek (2010))
This is a prerequisite for cell expansion. In the shoots of deepwater rice, submerged internodes accumulate considerably more expansins than those in air. The proteins occur in the intercalary meri- stem and in the adjacent extension zone but not in the differentiation zone (Cho and Kende 1997). Auxin also plays a role in shoot expansion: it stimulates acidification of the cell wall by activating the ATP-dependent proton pumps in the plasma membrane.
Conversely, when the SUB1A gene is present in rice, the GA response is inhibited through the activation of negative regulators. SLENDER RICE 1 and a related gene in rice encode DELLA proteins, which repress the induction of GA response genes such as expansins. The energy conservation is further supported by reducing the mobilisation of carbohydrate reserves—for instance, through the inhibition of sucrose synthase activity. SUB1A also supports recovery from post-anoxic shock upon subsidence of floodwater by up-regulating antioxidant defences. The function of SUB1A as a major regulator of flooding responses has enabled successful breeding approaches.
The differential developmental and metabolic response to ethylene is not unique for rice but conserved among flooding-tolerant plants. Species of the genus Rumex, as well as other plant species in the Rhine Valley floodplain, display varying degrees of flooding tolerance, and their growth behaviour can be placed at different positions along the continuum between escape and quiescence (Fig. 5.5). R. palustris tries to reach the atmosphere by hyponastic leaf growth (Fig. 5.10); the closely related species R. acetosa does not. Ethylene invariably accumulates in submerged tissues, yet only R. palustris shows activation of cell expansion.
These differences can also be seen when more species are tested. Ethylene treatment stimulates growth in only a subset of the species (Fig. 5.17). All of them—for example, Ranunculus sceleratus and Rumex maritimus (Fig. 5.5)—belong to the same niches in the Rhine Valley. These niches are defined mostly by gradients in flooding duration and the speed of soil drying after a flooding event. The escape strategy is associated with long-lasting floods and slow drainage.
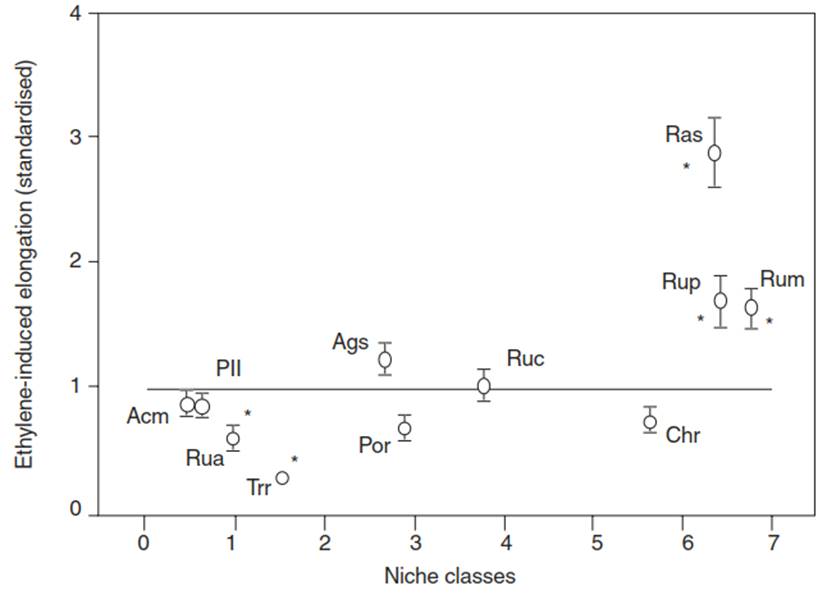
Fig. 5.17. Differential responses to ethylene in herbaceous species from floodplains of the Rhine River. For 11 species belonging to different niche classes, ethylene-induced shoot elongation (5-day treatment with air containing 10 μL/L ethylene) was determined relative to the mean elongation under control conditions (treatment with air) (standardised at 1). Asterisks indicate significant differences (p < 0.05) between ethylene and control treatments. The niche classes are characterised by differences in the duration of flooding and in the speed of run-off and drying after a flooding event. The elongating species are found in sites with long-lasting and slowly receding floods. Abbreviated species names: Acm, Achillea millefolium; Ags, Agrostis stolonifera; Chr, Chenopodium rubrum; Pll, Plantago lanceolata; Por, Potentilla reptans; Ras, Ranunculus sceleratus; Rua, Rumex acetosa; Ruc, Rumex crispus; Rum, Rumex maritimus; Rup, Rumex palustris; Trr, Trifolium repens. (Modified from Voesenek et al. (2004)
According to genome-wide transcriptome analyses of the two model Rumex species R. palustris and R. acetosa, using RNA sequencing, the differential behaviour can be associated with the activation of different processes similar to what has been found in rice. Submerged R. acetosa undergoes metabolic reprogramming consistent with quiescence, while R. palustris activates photomorphogenesis and shade avoidance pathways to support the growth response (i.e. escape).
References:
Amberger A. Pflanzenernahrung, 3rd edn, Ulmer. 1988.
Bailey-Serres J, Fukao T, Gibbs DJ et al (2012a) Making sense of low oxygen sensing. Trends Plant Sci 17: 129-138.
Bailey-Serres J, Lee SC, Brinton E (2012b) Waterproofing crops: effective flooding survival strategies. Plant Physiol 160:1698-1709.
Bailey-Serres J, Voesenek LA (2010) Life in the balance: a signaling network controlling survival of flooding. Curr Opin Plant Biol 13:489-494.
Bailey-Serres J, Voesenek LACJ (2008) Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 59:313-339.
Blom C, Voesenek L (1996) Flooding: the survival strategies of plants. Trends Ecol Evol 11:290-295.
Blom CWPM, Voesenek LACJ, van der Sman AJM (1993) Responses to total submergence in tolerant and intolerant riverside species. In: Jackson MB, Black CR (eds) Interacting stresses on plants in a changing climate, NATO ASI Series 1, vol 16. Springer, Berlin.
Cho HT, Kende H (1997) Expansins in deepwater rice internodes. Plant Physiol 113:1137-1143.
Christopher ME, Good AG (1996) Characterization of hypoxically inducible lactate dehydrogenase in maize. Plant Physiol 112:1015-1022
Date added: 2025-01-18; views: 579;