Plant Structural Diversity and Biodiversity
Looking at a plant community it becomes obvious that the species present may build up different structures, that is, different physical patterns. Vegetation structure is not only an important driver of the diversity of an ecosystem; it is also the key aspect that couples plants to the atmosphere, especially for gas exchange, trace gas fluxes, atmospheric deposition and rainfall interception.
Important structural characteristics of a plant include architectural traits, for example, height and branching patterns, leaf form and root stratification. Several attempts were made by plant geographers, as well as by ecologically orientated botanists, to differentiate between life forms based predominantly on structural features. Even the Greek philosopher Theophrastus (ca. 371— 287 BC) already thought about criteria by which to categorise plants according to their physiognomy, that is, the outer appearance of living things. He came up with a tree, shrub, perennial herb and “useful” plant categorisation. These attempts show that it is very difficult to cope with the great diversity of structural features in the plant kingdom (Table 20.3).
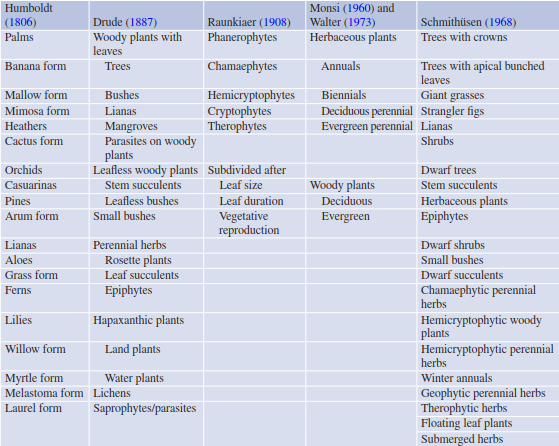
Table 20.3. Categories of plants by life form
In closed stands, the presence of strata of different heights caused by different life forms, intertwining with lianas, as well as standing or lying dead woody debris, increases structural diversity (Fig. 20.5). In addition, the architecture of the canopy can differ widely depending on the different growth forms of the plants. Such structures, in turn, provide habitat for many other organisms like birds, insects, epiphytes and lichens. According to the resource heterogeneity hypothesis and niche theory (Huston 1979) (Sect. 19.3), a larger variety of resources provide niches for other organisms with distinct requirements for habitat, food or environmental conditions.


Fig. 20.5. Structural diversity and species richness. A tropical rain forest in Indonesia a, rich in species and in structures, has a structural diversity that is similar to that of an extra-tropical rain forest b, poor in species but rich in structure. However, the number of higher plant species differs ten-fold between both forest types. Cultivated cereal fields are single layered and as such poorer in structure than forests. Nevertheless, there are large differences in species richness among differently cultivated fields: a species-rich but structure-poor fallow c and a species- and structurepoor cereal field d, both located in Mediterranean areas of Morocco
Hence, the structural heterogeneity or diversity of plant communities is often positively related to the biodiversity of other taxonomic groups, as shown by the classic example by MacArthur and MacArthur (1961) from temperate, deciduous forests of northeastern North America (Fig. 20.6, also Fig. 20.14). Therefore, maintaining or enhancing certain aspects of structural diversity is relevant for many aspects of nature conservation and management. For example, in retention forestry, single trees, decaying logs or small patches of trees are left at the time of harvest and are not removed in future management operations. The aim is to promote the maintenance of biodiversity and ecological functions, based on the rationale that biological structures and complexity beget niches and habitats and, thus, biodiversity.
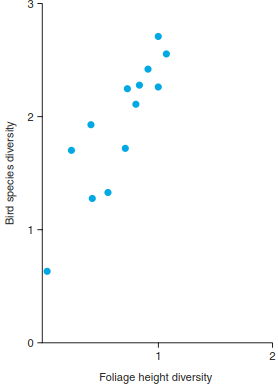
Fig. 20.6. Structural diversity in plant communities often begets biodiversity of other organisms. Bird diversity in temperate, deciduous forests of north-eastern North America is positively correlated with the structural diversity of the tree canopy. Foliage height diversity is a measure of habitat structural complexity and has been calculated as Shannon’s diversity index with the proportion of the total foliage in several horizontal layers. Bird species diversity is Shannon’s diversity index of breeding bird species territories recorded per 5 acres (approx. 2 ha). Two data points from a tropical savanna and from a pure spruce forest are also included
The structure of a plant community is also very relevant for the coupling between the biosphere and the atmosphere: the roughness of a canopy largely determines the exchange of water vapour and CO2 (Sects. 16.1 and 16.2) and strongly influences the microclimate (Sect. 9.2). For example, the daytime surface temperature of a forest near the alpine tree line is much lower than that of adjacent alpine grassland because the tall trees are coupled to the lower atmospheric layer, causing a transport of convective heat (Sect. 9.1). In contrast, small grasses and herbs are decoupled from atmospheric conditions, leading to a strong warming of the grassland canopy and top soil during periods of high radiation (Körner 2003).
In landscape ecology, where the importance of scale is in focus, biodiversity and the diversity of abiotic conditions or environmental heterogeneity are summarised as landscape diversity. Abiotic features of landscapes and the spatial heterogeneity and arrangement of structures are very important for biodiversity, and landscapes differ in biodiversity. It can be assumed that landscapes with spatially heterogeneous abiotic conditions may provide a larger variety of potential niches than those with more homogeneous conditions. For example, Schulze et al. (1996) showed that the richness and diversity of trees and shrubs were significantly higher at sites with high geo- morphological heterogeneity than at sites that exhibited little change in terrain or soil conditions in deciduous forest ecosystems. At the landscape scale, human-created structures must also be considered, for example, fragments of almost natural stands or linear elements such as hedgerows and windbreaks. At this level it is clear that structural heterogeneity contributes to high diversity and that diversity changes greatly in transitional areas of different ecosystems (ecotones), for species as well as for structures.
Date added: 2026-04-26; views: 73;